
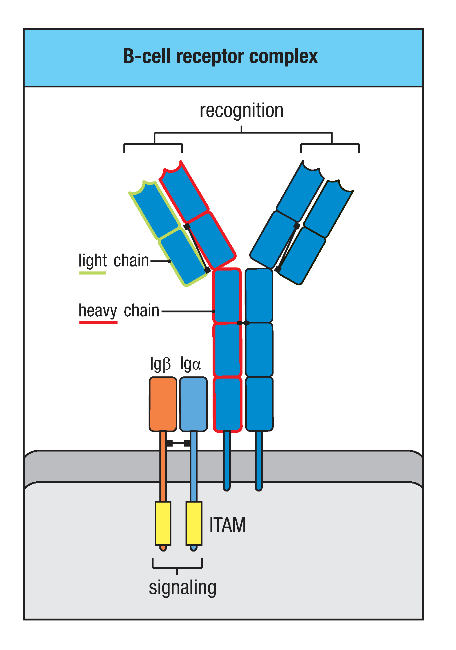
图1 BCR复合物结构
B细胞抗原受体(B-cell receptor, BCR)是一种位于B细胞表面的负责特异性识别及结合抗原的分子,其本质是一种膜型免疫球蛋白(mIg)。BCR具有抗原结合特异性,每个个体的BCR多样性高达5×1013,构成容量巨大的BCR库,赋予个体识别各种抗原,产生特异性抗体的巨大潜能。
BCR是由两条相同的分子量较小的轻链(L链)和两条相同的分子量较大的重链(H链)组成的四聚体结构。同分泌形式的Ig不同,BCR的重链是穿膜的多肽链。由于BCR的胞质区很短,不能直接将抗原刺激信号传递到B细胞内,因此完成BCR信号转导功能还需要Igα(CD79a)和Igβ(CD79b)的协助。Igα和Igβ都是免疫球蛋白超家族结构相关基因的表达产物,其在物种之间高度保守。Igα和Igβ都属于I型跨膜糖蛋白,在B细胞中以二硫键相连形成异源二聚体,并同mIg相连接。Igα、Igβ和mIg的跨膜区均有极性氨基酸,能够借助静电吸引而组成稳定的BCR复合物。Igα和Igβ的胞质区含有免疫受体酪氨酸激活基序(ITAM),能够募集下游信号分子,从而转导抗原与BCR结合所产生的信号。
BCR识别胸腺依赖性抗原(TD抗原)对B细胞激活有两个相关联的作用:(1)BCR可特异性结合抗原,产生B细胞活化的第一信号;(2)B细胞通过内化BCR所结合的抗原,并对抗原进行加工,形成抗原肽——MHCII类分子复合物(pMHC II),提呈给抗原特异性辅助T淋巴细胞(Th细胞)识别并使其活化。Th细胞活化后通过表达CD40L与B细胞表面的CD40结合,提供B细胞活化的第二信号。
蛋白激酶对酶和其他蛋白质的磷酸化是细胞调节其生化活性的普遍机制,具有调控迅速、反应可逆等优势。其中一个重要效应为募集胞质蛋白参与信号通路。例如,许多涉及信号转导的激酶是位于细胞膜内表面的,不能有效激活胞质中的靶蛋白;当受体活化,磷酸化的转接蛋白可以形成靶蛋白结合位点,招募细胞质中游离靶蛋白在激酶附近富集,继而依次被磷酸活化。
在抗原受体的信号转导过程中,由酪氨酸激酶反应产生的磷酸酪氨酸可以形成以一个蛋白质结合位点,即SH2结构域。这种结合位点在多种胞内信号转导蛋白质中发现,包括Src家族激酶。
1. BCR-Igα、Igβ介导的胞内信号转导
BCR在与抗原特异性结合后即启动B细胞活化的第一信号——ITAM上的酪氨酸磷酸化。B细胞内负责磷酸化ITAM的激酶有Blk、Fyn、Lyn三种,它们属于Src家族酪氨酸磷酸激酶。Src家族激酶是脊椎动物和其他动物中控制细胞分裂与分化相关的信号转导通路的共同成员。Src家族激酶可用过N端结构域与未磷酸化的ITAM低亲和力的结合,当抗原结合导致受体聚集时,受体相关激酶发生磷酸化并互相激活,使得Igα、Igβ尾部ITAM依次磷酸化。ITAM单个酪氨酸磷酸化可以使得其与Src家族激酶的SH2结构域结合,进一步磷酸化ITAM其他残基。Src家族激酶酶活性是由激酶结构域和C端区域的自我磷酸化调节的,这些区域均有调节性的酪氨酸残基。激酶结构中的酪氨酸发生磷酸化使其具有活性,在C端的酪氨酸发生磷酸化则抑制其活性。CD45可解除此抑制效应,是受体介导信号通路所必需的一步。
磷酸化的ITAM招募酪氨酸激酶(Syk)。Syk是BCR信号通路的关键激酶,含有两个SH2结构域,只有与B细胞受体的双重磷酸化ITAM结合之后才有酶活。
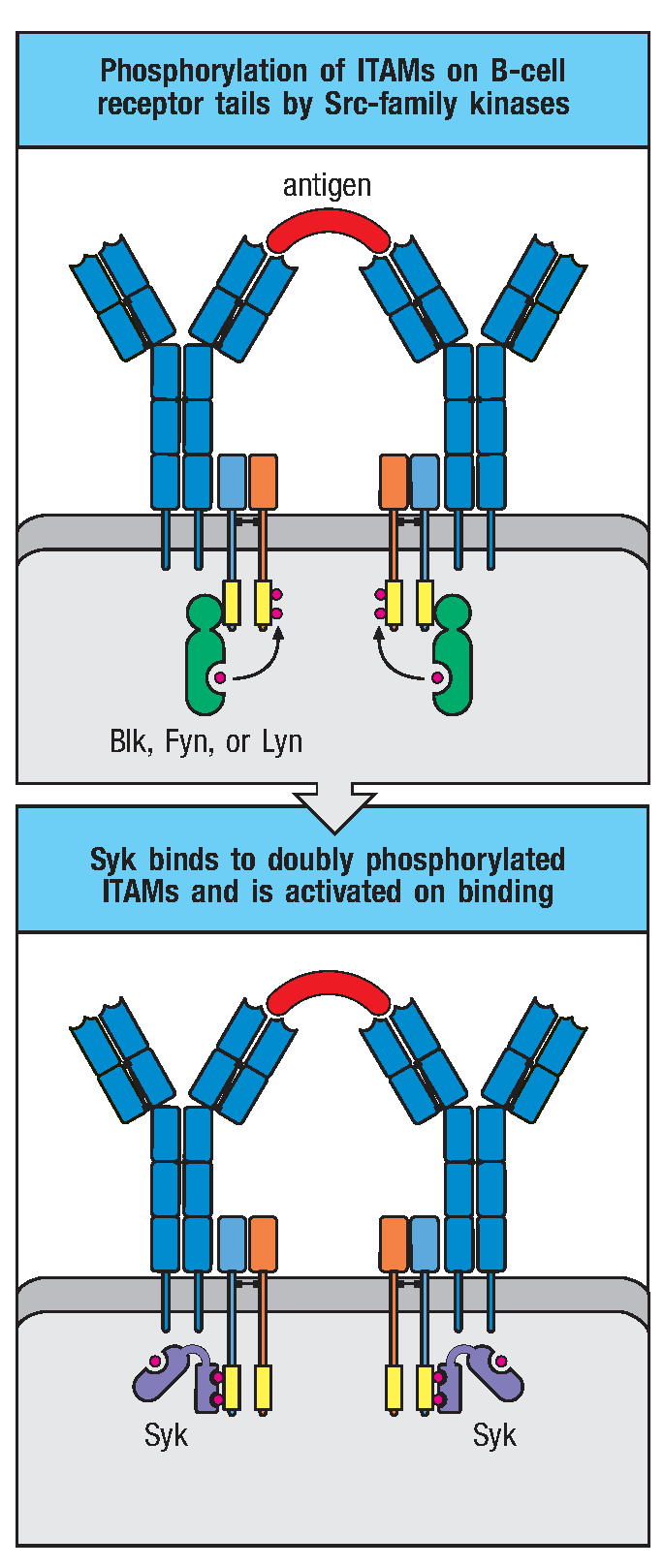
图2 BCR介导的胞内信号传导
Syk活化后使其他靶分子磷酸化,包括转接蛋白BLNK,由此招募Tec激酶,后者进一步激活磷脂酶Cγ2(PLC-γ2),催化细胞质内的磷脂酰肌醇二磷酸(PIP2)分解为三磷酸肌醇(IP3)和甘油二脂(DAG)。这两个分子都是信号通路中的重要组分。
IP3与内质网上的相应受体相互作用可引起Ca2+进入胞浆,内质网内腔的Ca2+浓度降低导致内质网膜蛋白STIM1与质膜上的钙离子通道ORAI1结合,促使ORAI1打开并让细胞外液的Ca2+流入细胞质,使信号得以维持。当细胞质中钙离子浓度增加时,会激活钙调蛋白(Calmodulin,CaM)使其构象改变。钙调蛋白可结合钙调神经磷酸酶(Calcineurin,CaN),CaN可以将NFAT(一种由钙离子通路调控的蛋白质,称为活化T细胞转录因子)上的磷酸化修饰水解,从而使其进入细胞核内启动下游基因转录。
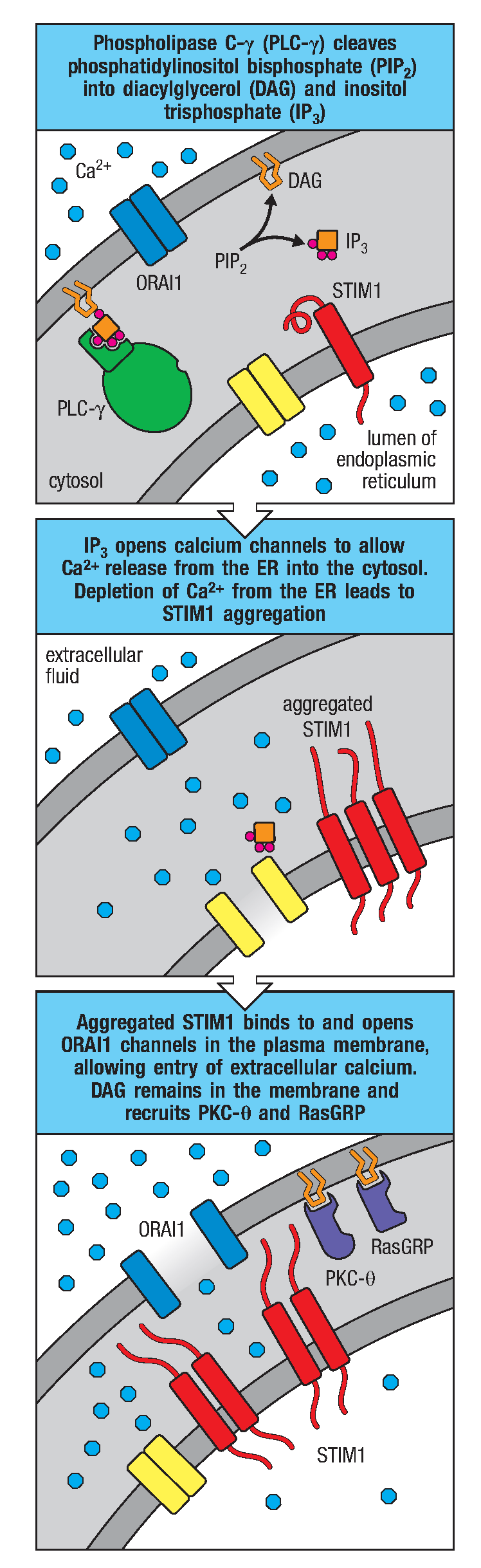
图3 钙离子信号通路
PIP2裂解的另一个产物为DAG,它可以保持与质膜内表面相联的状态,有助于活化蛋白激酶C(Protein kinaseC,PKC)家族成员。PKC家族大多为Ser/Thr蛋白激酶,可帮助启动通往细胞核的信号转导通路。另外,DAG也可招募RasGRP蛋白,RasGRP是Ras的一个鸟苷交换因子,含有C1结构域,用来激活附着于质膜上的Ras蛋白。Ras被激活后,下游再依次激活Raf、Mek、Erk三个激酶。因为Erk属于MAPK家族蛋白(mitogen-activated protein kinase),所以Ras激活产生的信号可传导到MAPK,MAPK再进入细胞核,磷酸化对应的转录因子。
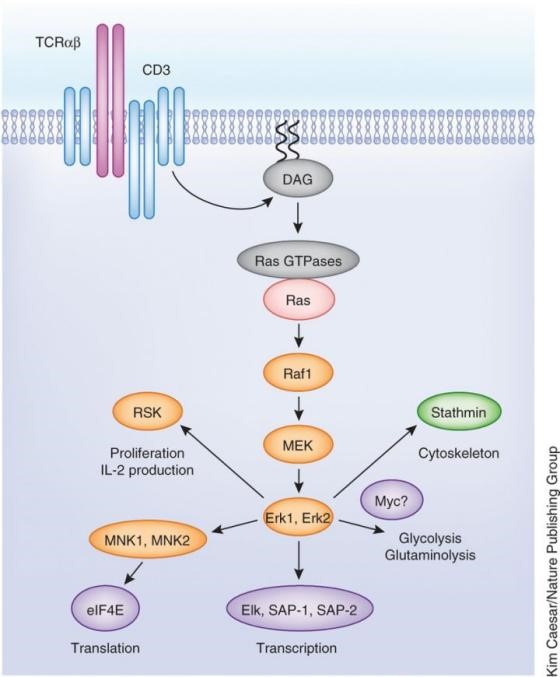
图4 DAG信号通路
最后通过蛋白激酶C(PKC)、丝裂原活化蛋白激酶(MAPK)及钙调蛋白等信号转导通路继续转导并最终激活NF-κB、AP-1、NFAT等转录因子,从而启动与B细胞活化、增殖、分化相关基因的表达。
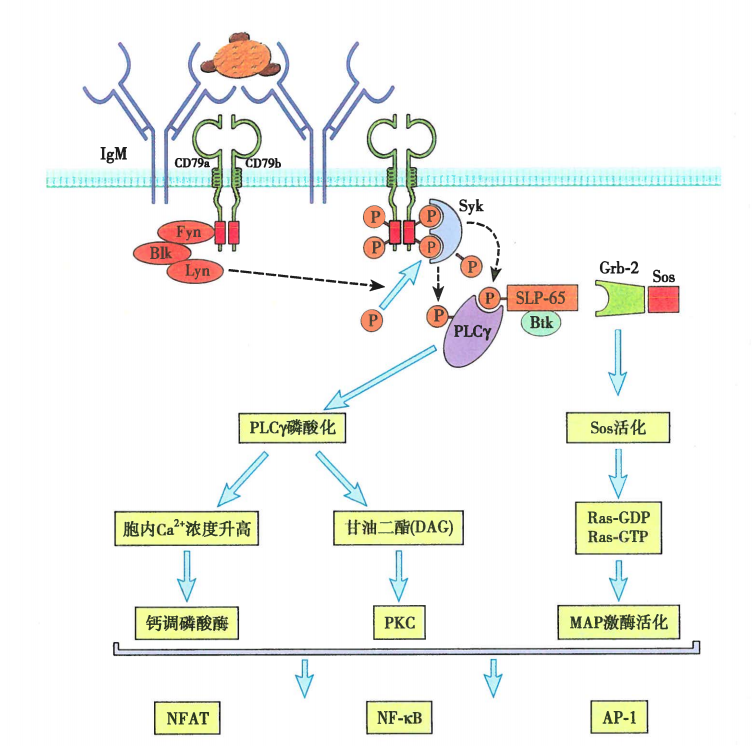
图5 BCR复合物介导的胞内信号转导
2. BCR共受体加强活化信号
B细胞表面的CD19/CD21/CD81复合物被称为B细胞共受体(B cell co-receptor),由CD19、CD21和CD81通过非共价键交联而成。复合物中,CD21可以和抗原结合从而增强BCR与抗原结合的稳定性,同时能识别与BCR-抗原结合的C3d,并通过交联CD19向胞内传递信号;CD19的胞浆区有多个保守的酪氨酸残基可以招募含SH2结构域的信号分子,与Igα/Igβ共同传递B细胞活化信号;CD81的主要作用可能是连接CD19和CD21,稳定B细胞共受体复合物。B细胞共受体通过稳定BCR与抗原的结合以及加强信号的传导,能够显著提高B细胞对抗原刺激的敏感性。
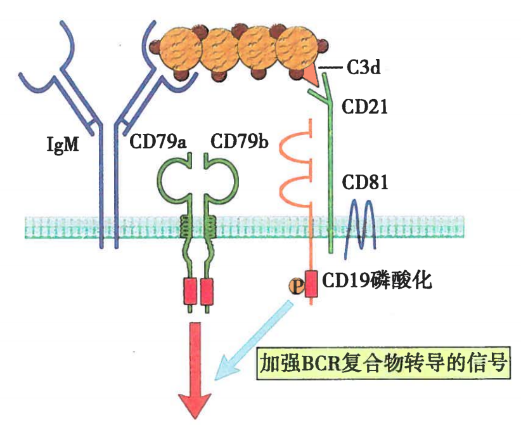
图6 B细胞共受体在B细胞活化中的作用
B细胞的第二活化信号又称共刺激信号,由Th细胞与B细胞表面多对共刺激分子相互作用产生,其中最重要的是CD40/CD40L。CD40表达在B细胞表面,CD40L表达在活化的Th细胞表面。CD40L与CD40相互作用,向B细胞传递活化的第二信号。
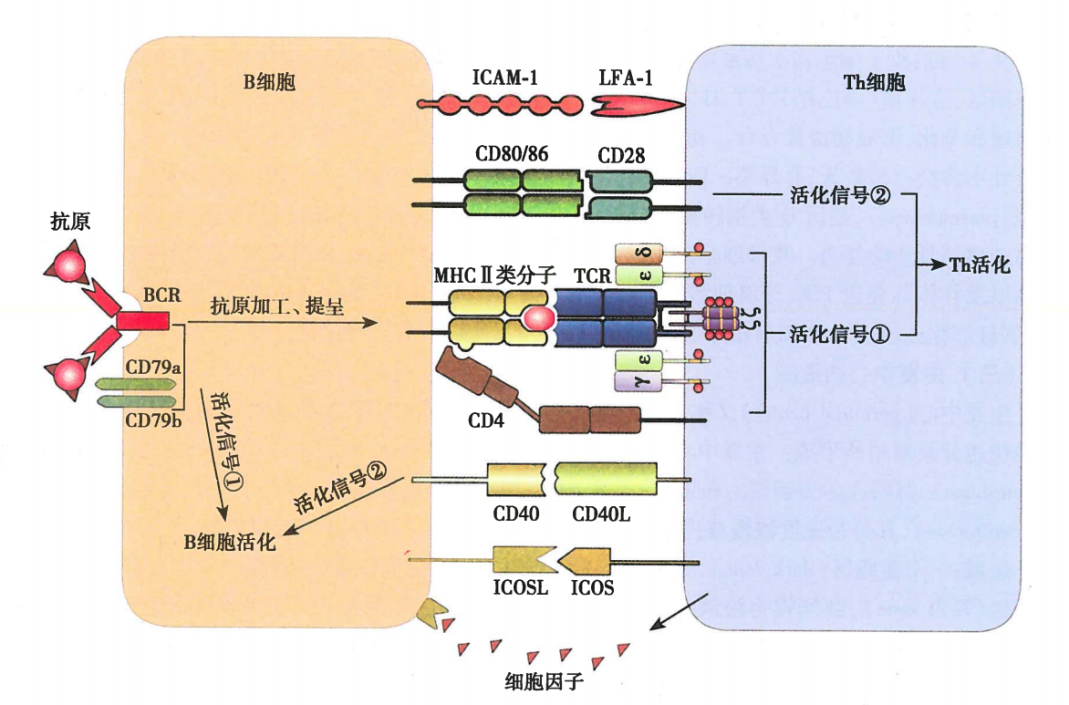
图7 B细胞与Th细胞相互作用
免疫球蛋白A(IgA)是呼吸道、胃肠道等黏膜表面分泌物中的主要抗体类别,由IgM通过抗体类别转换而来。IgA具有抑制细菌和病毒与上皮细胞的粘附,中和细胞内外病毒和细菌毒素的功能,被认为是抵御许多入侵病原体的重要第一道防线。IgA也是一种较弱的补体激活剂,在抗原有限的情况下,通过阻断强激活补体剂IgG或IgM抗体的结合抑制补体激活。
与其它免疫球蛋白(Ig)相同,IgA单体也由2条相同的重链和轻链组成,顶端2个Fab区可识别抗原,Fc区可与各种受体和效应分子相互作用。Fab区和Fc区由一个灵活的铰链区隔开。与其他Ig不同,IgA重链为α链,CH3包含一个称为尾部(Tp)的18个氨基酸C末端延伸。
在人体内IgA存在IgA1和IgA2两个亚类,IgA1是全身免疫系统产生的主要IgA亚类,IgA2主要存在于结肠。IgA1、IgA2重链恒定区(即Cα1、铰链、Cα2和Cα3)铰链序列差异显著(图1)。IgA1包含一个重复的八肽序列,范围大易水解;IgA2敏感铰链区序列缺失,抗蛋白酶水解能力更强。IgA2包含IgA2m(1)和IgA2m(2)两种同种异型,IgA2m(2)的H、L链通过二硫键连接,结构稳定;IgA2m(1)通过非共价相互作用,结构松散易分离。分子结构显示IgA1的2个Fab顶端间距比IgA2m(1)更大(图2),为此IgA1可能对表面具有重复抗原结构的病原体有更高的亲和力。

图1 人IgA1和IgA2铰链区比较

图2 人IgA1和IgA2m(1)分子模型
注:IgA1、IgA2m(1)轻链为绿色;IgA1重链为紫色,IgA2m(1)重链为青色。(图片来源于参考文献[3])
IgA在体内有分泌型IgA(SIgA)和血清IgA两种,分别来源于分布于黏膜相关淋巴组织的浆细胞和骨髓及外迁至脾中的浆细胞。分泌型IgA(SIgA)存在于唾液、眼泪、初乳、肠道、生殖道以及呼吸道的分泌物中,大多以聚合物(pIgA)形式存在,主要由二聚体(dIgA)组成。分泌型IgA(SIgA)分子量为385 kD,平均血清浓度为0.05 mg/mL。pIgA由IgA单体Fc区与J链尾部倒数第二个Cys残基通过二硫键连接而成。与单体相比,pIgA含有多个Fab,具有更强的抗原亲和力,更易于凝集外部分泌物中的微生物。
血清IgA直接分泌到血液中,以单体形式存在,其中约90%是在骨髓中产生的IgA1单体。血清IgA分子量为160 kD左右,占总Ig的10%–15%,浓度仅次于IgG(12 mg/mL)约为2-3 mg/mL,产生速率与IgG相似,但代谢速度比IgG快五倍。

图3 人IgA1、IgA2m(1)、IgA2m(2)、dIgA1以及SIgA1示意图
注:重链恒定区为红色,可变区为粉红色。轻链恒定区为蓝色,可变区为淡蓝色。J链为黄色,分泌成分为紫色。在IgA的单体形式上,O-糖基化为绿色(在IgA1铰链上),N-糖基化为深蓝色。(图片来源于参考文献[3])
pIgR介导的IgA黏膜上皮转运及功能
IgA由黏膜上皮固有层中的局部浆细胞(PC)产生。pIgA通过J链与聚合免疫球蛋白受体(pIgR)在基底外侧表面结合,形成的pIgR-IgA复合物被内吞(图4-③)并通过一系列囊泡穿过细胞被传递到顶端表面。在顶端表面的pIgR蛋白水解,释放SC-pIgA复合物(SIgA)或游离分泌成分(SC)(图4-④)。SC是一种亲水性和高度糖基化带负电荷的分子,在腔分泌物中,IgA与SC的共价偶联可以提升SIgA的亲粘特性和抗蛋白酶水解的稳定性。
转运过程中pIgA可以结合、中和和消除病毒。浸润的抗原和病原体被pIgA转运回管腔。亚上皮树突状细胞(DC)可以呈递抗原或吸收通过微皱褶细胞(M细胞)进入的SIgA包被的病原体。固有层中的病原体被dIgA包被形成免疫复合物,由表达FcαRI的DC和中性粒细胞摄取。中性粒细胞分泌白三烯B4(LTB4),吸引更多的中性粒细胞,从而清除感染。

图4 聚合免疫球蛋白受体(pIgR)介导的IgA黏膜上皮转运示意图
注:图中为上皮的简化横截面,顶部和底部分别具有顶端和基底外侧表面。pIgR为橙色,聚合IgA为红色,细胞核(N)和内质网为灰色,细菌病原体为深蓝色和粉红色,病毒病原体为黑色菱形。BEE为基底外侧早期核内体;CE为共核内体;ARE为顶端再循环内体;AEE为顶端早期内体。(图片来源于参考文献[3])
肠道系统中的SIgA功能
SIgA在维持对无害共生物的耐受及病原体免疫力平衡方面具有重要作用。在肠道系统中,SIgA从上皮层释放到管腔,存在于外粘液层中,SIgA涂层发挥分子“油漆”作用,干扰细菌运动并包围具有亲水外壳的病原体,阻止病原体进入肠上皮。SIgA通过抗体介导的表面抗原交联会凝集细菌,中和酶、毒素等细菌产物,再通过肠蠕动去除细菌聚集体。SIgA与粘液层形成抵抗病原微生物的天然屏障,共同防止病原微生物在黏膜上皮定植和渗透,避免感染。
FcαRI介导的IgA功能
血清IgA免疫复合物通过亲和力结合并交联FcαRI激活免疫细胞,从而诱导促炎反应,消除病原体。单体IgA通过可能IgAFc受体(FcαRI;CD89)抑制ITAMi信号传导维持稳态。由于pIgR相互作用位点与FcαRI部分重叠,SIgA与FcαRI结合受阻,无法通过FcαRI促进吞噬作用,但SIgA能够通过FcαRI和凝集素Mac-1(CD11b/CD18)的共同刺激呼吸爆发。与IgG或IgM相比,IgA在从血液中清除病毒的作用很小,并且对于再次接触病毒后对全身性病毒的保护性免疫是非必需的。
参考文献
[1] Sterlin D, Mathian A, Miyara M, et al. IgA dominates the early neutralizing antibody response to SARS-CoV-2[J]. Sci Transl Med, 2021, 13(577):eabd2223.
[2] Wang Z, Lorenzi JCC, Muecksch F,et al. Enhanced SARS-CoV-2 neutralization by dimeric IgA[J]. Sci Transl Med, 2021, 13(577):eabf1555.
[3] Macpherson, A., McCoy, K., Johansen, FE., et al. The immune geography of IgA induction and function[J]. Mucosal Immunol, 2008, 1(1):11–22.
[4] Breedveld A, van Egmond M. IgA and FcαRI: Pathological Roles and Therapeutic Opportunities[J]. Front Immunol, 2019, 10: 553.
天然IgM抗体与NK细胞、DC细胞、肥大细胞以及巨噬细胞是先天免疫系统的一部分,是抵御入侵微生物和异常人类细胞的第一道防线。这种免疫反应涉及与特定的抗原基序结合,例如糖蛋白或糖脂上的特定糖类或类似脂多糖的重复结构,由胚系(即未突变)基因编码的IgM抗体识别。通常,这些天然IgM抗体与一系列相似的外来抗原的亲和力不高,然而通过与这些外来抗原之间10个(在五聚体中)或12个(在六聚体中)位点的结合,其间的亲合力(avidity)被扩增。IgM抗体在锚定补体、调理吞噬作用的强效能力使其对细菌和病毒的防御特别有效。
IgM抗体与IgG的效应机制也有所不同。IgG利用NK细胞的参与,产生抗体依赖性细胞介导的细胞毒性(ADCC)以及补体依赖性细胞毒性(CDC)。IgM不结合Fcγ受体,因此不表现出ADCC。然而IgM具有非常强的CDC活性,它们的六聚体或五聚体结构允许其与补体C1q高度亲和结合,因此IgM能够比IgG更好地锚定补体。Sharp等人最近使用相板冷冻电子显微镜进行的工作提供了一个详细的模型,说明了抗原结合时是如何发生构象变化从而启动补体锚定的,同时也更清晰地了解到IgM与C1q结合的区域,即启动补体级联所需的第一个蛋白复合物。当Fab区在细胞表面上结合抗原时,游离IgM的平面或盘状结构变为“蹲伏”或“吻合样”结构。由于Cμ2区域的柔韧性,抗原结合Fab区从Cμ3,Cμ4和尾部肽成的环平面中移出。这允许许多或所有Fab臂接触表面抗原,从而利用IgM的亲合力。其他效应机制,如抗体依赖性细胞吞噬作用,也与IgM的作用有关。

图1 IgG与IgM结合和激活补体示意图
已知IgM抗体与多个受体结合,下面主要讨论这三种受体的功能作用。

图2 已知结合IgM的受体的示意图
1. 多聚免疫球蛋白受体(pIgR)
pIgR是一种单跨膜膜蛋白,其结构可分为胞外结构域、跨膜结构域和胞内结构域,其N端在细胞外C端在细胞内。整个蛋白由约620个氨基酸构成的胞外结构域、23个氨基酸构成的跨膜结构域和103个氨基酸构成的胞内结构域组成。pIgR具有五个Ig样结构域(D1-D5),这些结构域糖基化水平高。铰链区存在于D1和D2之间以及D3和D4之间。
pIgR与含有J链的聚合IgM或IgA抗体结合后,可被内化并由核内体从基底表面运输至顶端表面。上皮细胞黏膜表面端区域包含一个蛋白水解敏感位点,当核内体移动到顶端时,在该位点发生水解,这导致与pIgR胞外域结合的聚合Ig释放。pIgR未与配体结合的情况下,运输至上皮细胞黏膜表面端时也会被蛋白酶水解,在顶端作为8 kDa片段释放。这种被分裂后释放的部分称作分泌成分(secretory component,SC)。含有J链的聚合免疫球蛋白(如IgA和IgM)通常与分泌成分一起存在于黏膜表面。
pIgR在黏膜上皮的基底层表达,在小肠和大肠中表达最高,在肺、胰腺、肾脏和子宫内膜等组织中也可见表达。
2. Fcα/μR
在筛选能够结合IgM的小鼠淋巴瘤细胞系受体的过程中,人们发现了Fcα/μ受体。该受体大小约为70 kDa,属于免疫球蛋白超家族,并且被广泛糖基化。Fcα/μ受体的主要在巨噬细胞和生发中心的滤泡树突状细胞表达,分别以中等和高度亲和力与IgM和IgA结合,介导IgM/IgA抗原复合物的内吞作用。表达Fcα/μ受体与pIgR一样,Fcα/μ受体主要通过与Cμ3和Cμ4中的决定簇与IgM相互作用。
Fcα/μ受体分布在淋巴组织,包括淋巴结和阑尾中,在包括肾脏和肠道在内的非淋巴组织中也广泛表达,在肺、肝和心肌中观察到较低的表达。Fcα/μ受体在肠道巨噬细胞、浆细胞和潘氏细胞上的表达暗示了其在黏膜免疫的局部和全身作用。
3. FcμR(FAIM3/TOSO受体)
最新发现的与IgM相互作用的受体是FcμR,它是一种约60 kDa的跨膜唾液酸蛋白。FcμR也称为TOSO受体,在慢性淋巴细胞白血病B细胞上高度表达,并且已被证实在IgM结合时内化。它与pIgR和Fcα/μ的不同之处在于它只与IgM结合而不与IgA结合。
识别IgM的FcμR的CDR1区域可能非常短,只有五个氨基酸。值得注意的是,FcμR不需要J链来结合IgM,其相互作用区域主要被认为是与Cμ3和Cμ4。
在人类和小鼠中,FcμR在B细胞上的表达最强,此外在小鼠的粒细胞、巨噬细胞和树突状细胞上也观察到低表达。在人体内,该受体在T细胞和NK细胞上表达,但在骨髓细胞上没有表达。
FcμR在稳态中具有多种功能:在发育中维持适当的IgM-BCR表达水平,防止自反应性B细胞(特别是B-1)的过度激活,并防止自身免疫抗体介导疾病的发展。在外来抗原暴露后,根据抗原剂量,FcμR可以通过与B细胞结合直接改变B细胞反应的强度,至少在人类中,也可以通过增强T细胞反应间接改变。事实上,FcμR表达的增强和抑制作用已在T非依赖性和T依赖性抗原的体液反应中被发现,这表明抗原类型、剂量和细胞亚群特异性等其他调节机制的可能性。然而,FcμR−/−小鼠表面IgM-BCR表达的失调仍有待充分考虑其潜在影响,它可能有助于理解在没有FcμR的情况下对免疫反应发展的影响。
参考文献
[1]Keyt BA, Baliga R, Sinclair AM, et al. Structure, Function, and Therapeutic Use of IgM Antibodies. Antibodies[J], 2020, 9(4): 53. doi:10.3390/antib9040053
[2]Blandino R, Baumgarth N. Secreted IgM: New tricks for an old molecule. J Leukoc Biol[J], 106(5):1021-1034. doi: 10.1002/JLB.3RI0519-161R
[3]Liu J, Wang Y, Xiong E, et al. Role of the IgM Fc Receptor in Immunity and Tolerance. Front Immunol. 2019 Mar 22;10:529. doi: 10.3389/fimmu.2019.00529
体液免疫应可能产生IgM、IgD、IgG、IgA和IgE同种型的免疫球蛋白,每种免疫球蛋白都拥有独特功能,能够介导宿主防御入侵的病原体。IgM是进化过程中最早出现的,在脊椎动物中最为保守的抗体,也是所有脊椎动物唯一共有的抗体。IgM最初是以B细胞受体(BCR)的形式在B细胞分化早期中表达产生。在免疫反应的后期,IgM以含有10个抗原结合位点和连接(J)链的可溶性五聚体的形式,或含有12个抗原结合位点且无J链的六聚体的形式由浆细胞分泌产生。五聚体或六聚体IgM的分子量分别约为900或1050 kDa。由于IgM的多价性质,其表现出比二价IgG更高的抗原亲合力(avidity)。
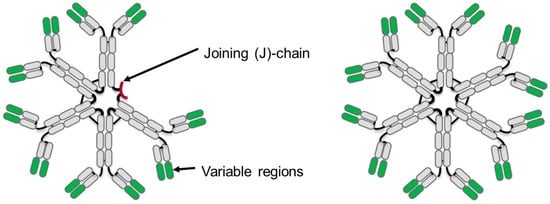
图1 IgM抗体五聚体(左)和六聚体(右)
注:恒定区域以灰色显示,可变区域以绿色显示,IgM五聚体上有以红色显示的J链。
IgM抗体通常以五聚体或六聚体的形式存在,其中每个单体约为190 kDa,由具有5个结构域(Vμ、Cμ1、Cμ2、Cμ3和Cμ4)的重μ链和具有2个结构域(Vκ-Cκ或Vλ-Cλ)的轻链组成。与其他同种型相比,IgM恒定区与IgE的同源性更大。IgG的重链恒定区CH1,CH2和CH3对应于IgM的Cμ1,Cμ3和Cμ4,IgG的铰链区对应于IgM的Cμ2,这是在其他同种型(哺乳动物IgE和禽类IgY)中发现也发现的额外恒定区。Cμ2的功能与IgG的铰链区域非常相似,并且提供了允许IgM在细胞表面上结合多个抗原所需的灵活性。每个单体中的重链在Cys 337处以二硫键共价连接。每个轻链都与重链中136位置的半胱氨酸残基结合。
μ链的另一个特征是在C端存在一个短的18个氨基酸肽序列(PTLYNVSLVMSDTAGTCY),称为尾部肽(tailpiece)。IgM单体通过这些尾部肽的倒数第二个半胱氨酸之间的二硫键共价连接。尾部肽对于IgM聚合至关重要。事实上,尾部肽在其它抗体同种型(如IgG)的C末端融合时,可以诱导聚合。此外,Cμ3中Cys 414残基之间的单体间二硫键将IgM良好的保持为环状结构。
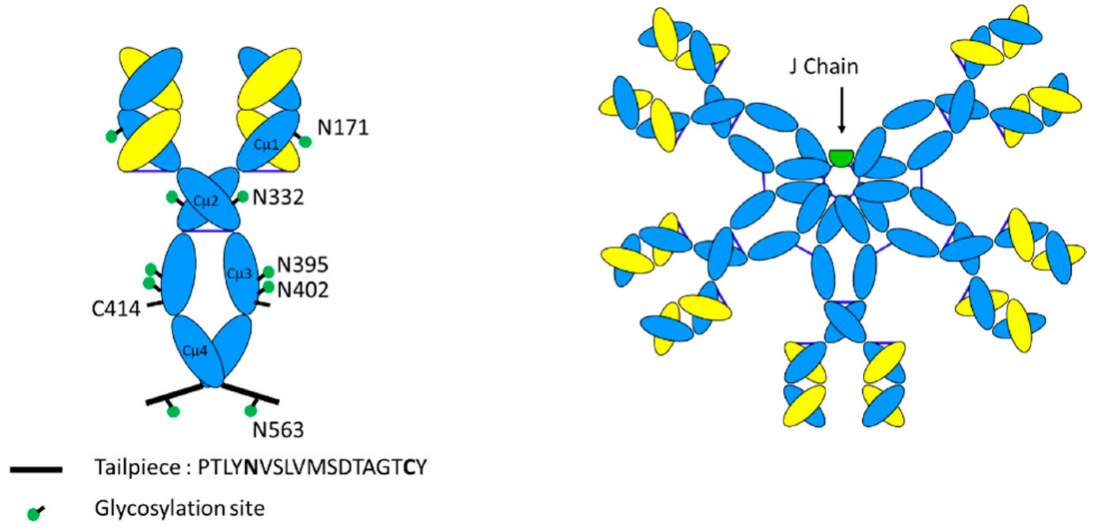
图2 IgM单体与IgM五聚体的关系示意图
注:IgM单体与IgG单体的区别在于它们在指示(绿色点)的天冬酰胺残基处的广泛糖基化,存在额外的结构域Cμ2代替铰链,以及存在对多聚化至关重要的短尾部肽序列。五聚体IgM还具有额外的137个氨基酸组成的J链。
除重链和轻链外,IgM还拥有第三条链,即一种由137个氨基酸组成的多肽,称为连接(J)链,这是IgA和IgM抗体聚合的关键特征。J链是一个独立的结构域,与重链和轻链的蛋白质折叠无关。从两栖动物到人类,J链的序列高度保守,在功能上帮助IgM五聚体和IgA二聚体通过与多聚免疫球蛋白受体(pIgR)的相互作用结合,运输到膜表面,体现了关键结构与功能相统一观点。
抗体是具有N连接糖基化的糖蛋白。IgG在Asn 297处存在N-糖基化,影响其与Fc γ受体的结合,在调节抗体依赖性细胞毒性(ADCC)中起作用。值得注意的是,与IgG相比,IgM具有更多的糖基化位点。IgG重链只具有单个糖基化位点,但人和非人灵长类动物IgM重链在Asn 171(Cμ1),Asn 332(Cμ2),Asn 395,Asn 402(均在Cμ3中)和Asn 563(位于尾部肽中)表现出5个N-糖基化位点。另一个糖基化位点存在于Asn 49的J链上。这些糖基化被认为有利于IgM结构的聚合和组装,并为IgM提供更高的溶解度和更长的体内半衰期。暂没有证据证明IgM的糖基化与介导其他效应有关。
糖基化位点增加了IgM抗体的复杂性。对IgM具体位点特异性糖类分析表明,并非所有N-连接位点都具有相似的糖基化。N-糖基化具有各种复杂程度,从高甘露糖、简单聚糖到双触角、三触角和四触角复合聚糖。Asn 171,Asn 332和Asn 395的三个位点表现为具有唾液酸苷化末端的复杂糖类。Asn 402和Asn 563更多为羧基末端的高甘露糖结构。这种糖基化模式与IgM N端区域更容易被细胞内高尔基体的糖基化酶接近,而C端区域没有充分修饰可能是由于空间位阻使得聚糖与聚合形式IgM间缺乏可及性。IgM上的第四个糖基化位点(Asn 402)与IgG的单位点同源,已知其具有有限的可及性,在IgM或IgG中都未连接复杂糖类。
尽管已知μ链尾部肽和J链在IgM抗体的组装和功能中起着关键作用,但多年来,IgM五聚体的三维结构在很大程度上未有定论。事实上,直到最近对J链二级结构的认知才有所进展。在新的关键研究中,展示了IgM五聚体的冷冻电子显微镜(cryo-EM)结构,令人惊讶的是,IgM五聚体并没有像假设的那样形成五边形结构,而是以不规则的五边形形式,在五个IgM单体中的两个之间有一个 50°角的间隙。
后有研究发现,这个间隙可以结合一种血清蛋白,即巨噬细胞表达的细胞凋亡抑制剂(AIM/CD5L)。因此,分泌型IgM不仅作为效应蛋白,而且还可以作为另一种效应蛋白的转运蛋白。
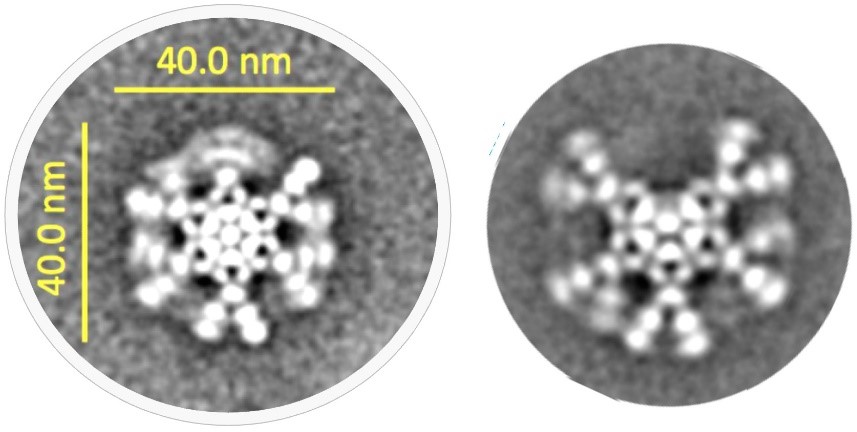
图3 抗CD20 IgM的六聚体(左)和五聚体(右)形式的冷冻电镜下的结构图
在哺乳动物中,B细胞发育在胚胎发育期间以层级、有序的方式在胎儿肝脏中发生,后在成人的骨髓和外周淋巴组织中发生。在骨髓中,CD34+多能祖细胞分化成常见的淋巴细胞前体(CLP),产生B细胞和T细胞谱系。CLP随后分化成表达Igα和Igβ的早期祖B细胞,Igα和Igβ是BCR信号传导的基本组成部分。
在向祖B细胞过渡期间,发生μ链(μHC)位点重排。RAG1/2重组复合物首先诱导D-JH重排,随后V-DJH重排。B细胞分化成large pre-B细胞之前,细胞表面不表达μHC,large pre-B细胞阶段,细胞表面表达BCR前体,由μHC复合物及替代轻链VpreB和λ5链组成。通过BCR前期的信号传导导致增殖、分化和随后的替代轻链下调,为λ或κ轻链重排,形成具有不同抗原特异性的表面IgM铺平了道路。受体编辑和选择发生在此时发生,表面表达IgM的未成熟B细胞从骨髓流出到脾脏。
在脾脏内,表达IgM的未成熟B细胞开始在细胞表面表达IgD,其分为以下不同的群体:在滤泡内的低表达IgM高表达IgD B细胞和在脾脏的边缘带的高表达IgM低表达IgD B细胞。B1细胞也成熟成高表达IgM低表达IgD细胞。尽管IgM和IgD的可变区相同,但选择性转录和剪接都会影响IgM和IgD重链。这些未成熟B细胞群现在已经成熟,并且在遇到抗原时可以进行克隆扩增和体细胞超突变。抗原结合后,通过IgM和/或IgD BCR启动信号传导,通过共受体CD19产生涉及Lyn、Syk、Src、Btk、PLCγ2和PI3Kδ的信号级联,导致产生分泌IgM、IgG、IgA或IgE的B细胞的活化、增殖和分化。而B1细胞仅产生分泌型IgM。
参考文献
Keyt BA, Baliga R, Sinclair AM, et al. Structure, Function, and Therapeutic Use of IgM Antibodies. Antibodies: 2020;9(4):53. doi:10.3390/antib9040053
抗体(Antibody,Ab)是介导体液免疫的重要效应分子,是免疫系统在外来颗粒或生物体的抗原刺激下,由B细胞或记忆B细胞增殖分化成的浆细胞所产生的、可与相应的抗原发生特异性结合的免疫球蛋白(Immunoglobulin,Ig),主要分布在血清中,也分布于组织液、外分泌液及某些细胞膜表面。Ab的基本结构是由两条完全相同的重链和两条完全相同的轻链通过二硫键连接呈的Y形单体,根据重链恒定区的差异可分为IgM、IgG、IgA、IgD和IgE五类同种型。
IgA与IgG简介
IgA是呼吸道、胃肠道等黏膜表面分泌物中的主要抗体类别,是机体抗原物质的主要来源。IgA具有抑制细菌、病毒与上皮细胞的黏附,中和细胞内外病毒和细菌毒素的功能,被认为是抵御病原体入侵的第一道防线。IgA有血清型和分泌型两种,血清IgA直接分泌到血液中,以单体形式存在,其中约90%是在骨髓中产生的IgA1单体。血清IgA分子量为160kD左右,占总Ig的10%–15%,浓度约为1.5~2.6mg/mL,仅次于IgG(9.5~12.5mg/mL),产生速率与IgG相似,但代谢速度比IgG快五倍。
分泌型IgA(SIgA)是外分泌液中的主要抗体类型,参与黏膜局部免疫,在局部抗感染中发挥重要作用,是机体抗感染的“边防军”。分泌型IgA(SIgA)主要以二聚体(dIgA)为主,在肠道、呼吸道、乳腺、唾液腺和泪腺等部位合成,经黏膜上皮细胞分泌至胃肠道和支气管分泌液、初乳、唾液和泪液中。SIgA通过特异性结合,阻止病原体黏附到细胞表面。
免疫球蛋白G(Immunoglobulin G, IgG)
IgG是二次免疫应答产生的主要同种型抗体,亲和力高、分布广泛,是机体抗感染的“主力军”。IgG在人出生后3个月开始合成,是血清和胞外液中含量最高的免疫球蛋白,约占血清总Ig的75%~80%,半衰期为20~23天,是半衰期最长的同种型Ig。基于重链恒定区,特别是CH1和CH3的结构、抗原和功能差异,IgG分为IgG1、IgG2、IgG3、IgG4四个亚类。二次免疫反应中,抗原诱导产生IgG亚型因抗原类型而异,蛋白质抗原通常诱导产生IgG1和IgG3抗体,多糖类抗原通常诱导产生IgG2和IgG4抗体。
IgA、IgG等同种型抗体是通过IgM同种型类别转换产生的。B细胞在Ig重链V区基因重排后其子代细胞中的重链V区基因保持不变,但位于VDJ基因座下游的是九个功能性CH基因则会发生重排,即同种型类别转换。Cμ、Cδ、Cγ3、Cγ1、Cα1、Cγ2、Cγ4、Cε、Cα2九个CH基因,除了Cδ外,每个CH基因的前面都有一个不能翻译的外显子和一个开关区(S区)。在同种型转换期间,中心细胞的CH区经历一系列DNA切割或重新连接事件,一旦接收到转换信号,DNA很可能被环出,中间未使用的CH外显子将被删除,选定的CH外显子与重排的VDJ重链外显子连接。VDJ重链外显子与新选定的外显子剪接产生mRNA,该mRNA被翻译成新的重链,导致B细胞从IgM转换为新的Ig类型。
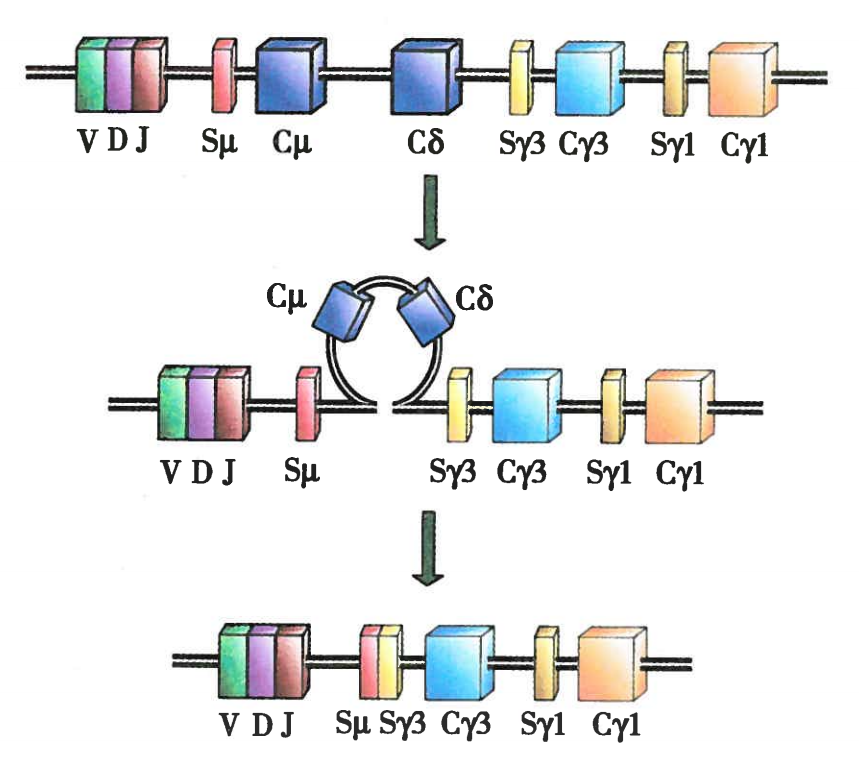
图1 Ig类别转换
注:Sμ、Sγ3、Sγ1分别为Cμ、Cγ3、Cγ1基因上游的转换区,在Ig类别转换为IgG3时,Sμ和、Sγ3重组,期间的序列包括Cμ、Cδ都被环出,从而使Ig类别转换为IgG3。
IgA和IgG之间的主要区别在于分泌多肽,IgA具有促进通过黏膜表面分泌的分泌多肽,而IgG没有。IgA具有J链(Joining chain)和分泌成分(Secretory component,SC)两种抗体辅助成分。J链是一种富含半胱氨酸的酸性糖蛋白,包含124个氨基酸,分子量约15kD,由浆细胞合成。J链通过尾部的倒数第二个Cys残基与单体IgA形成二硫键,聚合IgA。J链是IgA上皮细胞间的转运及其在黏膜表面分泌的先决条件,对IgA分泌至关重要。SC是多聚免疫球蛋白受体(pIgR)的胞外段,是一种由黏膜上皮细胞合成、分泌的含糖肽链,分子量约为75kD。多聚体IgA通过J链与pIgR结合,辅助SIgA由黏膜固有层,经黏膜上皮细胞转运,分泌到黏膜表面,最终pIgR蛋白水解并释放SC-pIgA复合物(SIgA),SIgA中的SC可保护其铰链区免受蛋白水解酶降解作用。
此外,单体IgA重链C端存在一个18个氨基酸组成的尾部,IgG重链中并没有这种结构。IgG、IgA抗体结合抗原表位的个数(即抗原结合价)也存在差异,单体IgG、IgA为2价,分泌型IgA(主要是二聚体IgA)为4价。
分泌型IgA可被转运到呼吸道和消化道黏膜表面,在黏膜局部免疫中发挥重要作用。黏膜上皮固有层中的局部浆细胞(PC)产生IgA。聚合IgA(主要是二聚体IgA)通过J链与聚合免疫球蛋白受体(pIgR)在基底外侧表面结合,形成的pIgR-IgA复合物被内吞并通过一系列囊泡穿过细胞被传递到顶端表面。在顶端表面的pIgR蛋白水解,释放SC-pIgA复合物(SIgA)或游离分泌成分(SC)。
IgG穿过胎盘的作用是一种重要的自然被动免疫机制。胎盘母体一侧的滋养层细胞表达一种IgG输送蛋白,称为新生Fc段受体(FcRn)。FcRn通过CH2-CH3结构域与IgG结合,介导母体IgG对新生儿的转胞吞作用,调节血清IgG水平。IgG与FcRn的结合严格依赖于pH,在体内酸性环境下与FcRn结合,通过胞吞作用转移到滋养层细胞内,并通过pH变化释放到进入胎儿血液循环中。FcRn结合可保护其免受溶酶体的破坏。
补体激活
补体级联激活是清除调理病原体的重要手段,抗体与相应抗原结合后,因构型改变而使其CH2和CH3结构域内的补体结合位点暴露,从而通过经典途径激活补体系统,产生多种补体的效应功能。IgG不同亚型补体激活能力不同,IgG4是唯一不能激活补体的IgG亚类,而IgG1、IgG2、IgG3对补体激活成分亲和力差异显著。IgG1和IgG3激活补体的能力较强,IgG2较弱,以C1q补体成分为例,IgG的亲和力大小依次为IgG3>IgG1>IgG2。相比于IgG,IgA是一种较弱的补体激活剂,可通过旁路途径激活补体系统。在抗原有限的情况下,通过阻断强补体激活剂IgG或IgM抗体的结合抑制补体激活。
婴幼儿被动免疫
人类初乳中分泌的IgA浓度非常高,含有针对多种微生物抗原的SIgA,婴儿可从母亲初乳中获得SIgA,在肠道中SIgA发挥分子“油漆”作用,干扰细菌运动并包围具有亲水外壳的病原体,阻止病原体进入肠上皮。SIgA通过抗体介导的表面抗原交联会凝集细菌,中和酶、毒素等细菌产物,再通过肠蠕动去除细菌聚集体。SIgA与黏液层形成抵抗病原微生物的天然屏障,共同防止病原微生物在黏膜上皮定植和渗透,避免感染。研究显示,母乳喂养可显著降低婴儿死于胃肠道和呼吸道感染的几率。
与IgA分子“油漆”作用不同,IgG可以穿过胎盘屏障,进入胎儿血液循环发挥被动免疫作用。IgG是唯一能通过胎盘的免疫球蛋白,对新生儿抗感染免疫至关重要。
表1 IgA、IgG主要理化性质及生物学功能
| 参数 | IgA | IgG |
|---|---|---|
| 开始合成时间 | 出生后4~6个月 | 出生后3个月 |
| 主要存在形式 | 单体/二聚体 | 单体 |
| 合成率[mg/(kg•d)] | 65 | 33 |
| 占总血清Ig比例 | 10~15% | 75~85% |
| 血清含量mg/mL | 1.5~2.6 | 9.5~12.5 |
| 半衰期(天) | 6 | 23 |
| 单体分子量(kD) | 160 | 150 |
| 重链 | α | γ |
| 亚类 | IgA1、IgA2 | IgG1、IgG2、IgG3、IgG4 |
| 抗原结合价 | 2,4 | 2 |
| 糖基化修饰 | 7% | 3% |
| 胎盘转运 | – | + |
| 结合SPA | – | + |
| 经典途径补体激活 | – | + |
| 旁路途径补体激活 | IgA1+ | IgG4+ |
| 主要作用 | 黏膜免疫 | 再次免疫应答 |
参考文献
[1]Schroeder HW Jr, Cavacini L. Structure and function of immunoglobulins[J]. J Allergy Clin Immunol., 2010,125(2 Suppl 2):S41-52.
[2]Macpherson, A., McCoy, K., Johansen, FE., et al. The immune geography of IgA induction and function[J].Mucosal Immunol, 2008: 11–22.
[3]医学免疫学[M].
多样化的抗体库对有效的体液免疫是必不可少的。体细胞超突变和转换重组在脊椎动物物种中广泛应用,体细胞基因转换作为一种免疫机制,在很大程度上限于在肠道相关淋巴组织(GALT)中初级抗体成分多样化的物种,例如兔子。这些物种通过一种与小鼠和人类完全不同的策略产生初级抗体库。免疫球蛋白(Ig)重链和轻链位点V(D)J基因重排过程中,小鼠和人类的B细胞通过利用许多不同的V(D)和J基因片段产生高度多样化的抗体库。而兔B细胞在V(D)J基因重排过程中,优先利用VH1基因,重排产生的V(D)J基因作为后续抗体库多样化的突变底物。重组的V(D)J基因通过体细胞基因转换和体细胞超突变引入突变,这一过程产生了大量不同特异性的抗体,可以用于预测和抵御大量潜在病原。
在兔的骨髓中,V(D)J基因重排产生初始抗体库,这一过程受B细胞优先使用VH1限制。兔子出生后约2天,B细胞离开骨髓,迁移到肠道相关淋巴组织(GALT),滤泡开始发育。GALT是幼兔B细胞增殖和V(D)J基因多样化的主要位点,最大的GALT就是阑尾。B细胞迁移似乎会持续约1-2周,此后会急剧下降。B细胞在阑尾滤泡顶端聚集,同时形成滤泡树突状细胞(FDC)网络。大约1周时,特定共生肠道微生物的定植密度和阑尾微环境的发育达到了足以启动B细胞增殖和初级抗体库多样化阶段。在滤泡基底区B细胞开始增殖,强健的B细胞增殖驱动快速的滤泡扩张,在此过程中B细胞广泛多样化其V(D)J基因。在断奶前后的发育期(3-4周龄时),母乳中提供的多样化抗体库开始减少时,初级抗体库的多样化开始,表达多种V(D)J基因的B细胞迁移到外周,在随后的3周内高度多样化的初级抗体库建立。
V(D)J基因重排是抗体库发育的关键,新生兔和成年兔B细胞中的大多数V(D)J基因后区都进行了有效的重排。VH区域是由三个非相邻基因VH、D和JH的重排在体细胞上产生的。兔子是已知的少数几种在Ig重链基因重排过程中有限使用VH、DH和JH基因片段组合连接的脊椎动物之一。V(D)J基因重排过程中小鼠和人可以利用很多VH,但对于兔子来说,80-90%的V(D)J基因优先采用VH基因片段VH1。剩余10-20%的V(D)J基因重排是采用另外两个VH基因片段VHx和VHy。兔子所有的VH基因都属于一个VH家族。尽管50-100种系VH基因具有功能,但可利用的VH基因数量有限,整个兔Ab基因库仅由4个VH基因产生。由于内含子调节或相对较低的甲基化程度等未知原因,VH1优先重排。在小鼠和人类中,VH基因的优先利用主要发生在胎儿期和新生儿期,成年期不明显。兔子VH1基因的优先利用发生在整个生命周期。
B细胞体细胞超突变(可能是非模板化的)和类基因转换(模板化的)事件的结合打破VH1基因优先使用的限制,实现了初级抗体(Ab)基因库多样性。虽然兔子VH区内的体细胞基因转换和超突变发生在相似的时间和位置,但兔子的抗体多样性大部分是由重组V(D)J基因的体细胞基因转换产生的。体细胞基因转换是将上游VH基因段供体的核苷酸序列转移到重排的VH基因段,通过CDR1和CDR2的不同组合产生组合位点多样性,这种机制随机性小,引入有害变化的概率低。另一种体细胞超突变可以将点突变引入到整个V(D)J基因,对高频率点突变进行靶向积累。值得注意的是,D区在重排的V(D)J基因中没有体细胞基因转换的上游供体基因,所以D区点突变只能通过体细胞超突变引入。
对于亲和力成熟来说,体细胞基因转换与超突变大有不同。通过体细胞基因转化可以快速产生多种B细胞受体。在Ag特异性免疫反应的背景下,体细胞基因转换(尤其是在CDR内)可能会改变Ag受体的特异性,而非亲和力。B淋巴细胞在成年兔中的生成是有限的,因此不会持续产生新的特异性Ag受体。体细胞基因转换与Ag诱导的外周受体编辑相似,可增加Ab库的整体多样性,挽救注定死亡的细胞。
产生和维持抗体多样性的机制因物种而异。在兔的T细胞依赖性免疫反应中,除了利用体细胞超突变使其IgH基因多样化外,还使用了类似体细胞基因转换的机制。小鼠和人的Ab多样性主要依靠V(D)J基因重排,Ig基因在生发中心通过体细胞超突变实现多样化,很少发现体细胞基因转换。绵羊胎儿和新生儿发育过程中的Ab基因多样性依赖于体细胞超突变而非基因转换。鸡的VH、VL基因片段多样化的主要机制是基因转换。
在通过体细胞多样化建立初级Ab库的物种中,如鸡、羊和牛等,组合重排对多样化的贡献很小,且它们的多样化发生独立于外源性因素,开始于出生前,受发育调节。兔初级Ab的体细胞多样化依赖于来自肠道的外源性因素(如肠道微生物),开始于出生后1-2个月,不受发育调控,且GALT和肠道菌群之间的相互作用是Ab基因多样性所必需的。GALT的充分发育需要特定种类的常规兔肠道菌群,更需要Ab菌群的多样化。肠道微生物可能是通过B细胞超抗原直接刺激滤泡B细胞增殖和V(D)J基因多样化,也可能是通过识别微生物高度保守的分子模式的toll样受体,触发激活途径,刺激广泛的B细胞增殖和V(D)J基因多样化。
参考文献
[1]Shi-Kang Zhai, Dennis K. Lanning.Diversification of the primary antibody repertoire begins during early follicle development in the rabbit appendix[J].Molecular Immunology, 2013,54:140-147.
[2]Lanning DK, Knight KL. Diversification of the primary antibody repertoire by AID-mediated gene conversion[J]. Results Probl Cell Differ, 2015,57: 279-293.
[3]Dennis Lanning, Periannan Sethupathi, Ki-Jong Rhee, et al. Intestinal microflora and diversification of the rabbit antibody repertoire[J].J Immunol, 2000,165 (4): 2012-2019.
[4]Candace R. Winstead, Shi-Kang Zhai, Periannan Sethupathi, et al. Antigen-induced somatic diversification of rabbit IgH genes: gene conversion and point mutation[J].J Immunol, 1999, 162(11): 6602–6612.
[5]Tunyaplin C, Knight KL. Fetal V(D)J gene repertoire in rabbit: evidence for preferential rearrangement of VH1[J]. Eur J Immunol, 1995, 25(9):2583-2587.
不同物种B淋巴细胞的发育途径不尽相同。小鼠和人类的B细胞的分化发育过程之前已有详细描述,B祖细胞会经历几个有序的发育阶段,这些阶段由V(D)J基因重排的状态和特定细胞表面标记物的表达与否来定义。在绝大多数哺乳动物中,B细胞的生成从胎儿肝脏开始,在胎儿末期转移到骨髓,并在骨髓中持续生成到成年。通常认为骨髓中B细胞的持续增殖是补充外周B细胞供应所必需的,从而可以在整个生命周期中维持多样的免疫活性B细胞库。然而与大多数哺乳动物不同,兔的B细胞生成期较短,且发生在个体发育早期。
兔B谱系淋巴细胞的分化最早在妊娠第14天的兔子中观察到。在兔子妊娠17~19天后,开始可以鉴定出胎肝中的pre B细胞。同时,胎儿网膜及脾脏中也存在pre B细胞。肝脏中pre B细胞的水平在出生时达到峰值,在出生后第10天几乎无法检测到。
与其他物种一样,兔体内的B细胞在胎儿末期开始从肝脏转移到骨髓。妊娠第25天,pre B细胞首次出现在胎儿骨髓中,出生后数量继续增加。出生至2周龄,兔骨髓中的pre B细胞逐渐达到峰值水平,占总有核细胞(total nucleated cells)的9%~19%。此后pre B细胞数量迅速减少,直到在成年兔骨髓中pre B细胞仅占总有核细胞的1%~3%。尚不清楚成年兔中的这一小部分前体细胞是否可以持续产生新生成的B细胞并进入外周免疫器官,或者它们在成年兔体内相对不活跃,且并非新成熟B细胞的重要来源。到16周龄时,骨髓中所有B谱系细胞基本都是B细胞。B细胞的增加可能是由于外周B细胞归巢回骨髓,而不是持续的B淋巴细胞生成。
兔B细胞的产生不会持续到成年。给新生IgH同种异型杂合的兔子注射针对父系IgH同种异型的抗体,会抑制该同种异型Ig的表达。这种抑制持续至少两年,甚至兔子的一生。这种抑制之所以是可持续的,是因为成年兔很少产生新生成的B细胞。这与在小鼠体内进行的同种异型抑制实验结果截然不同。在IgH抑制的小鼠中,该同种异型B细胞在6周内重新出现,这可能是B细胞增殖的结果。对这些新生成的B细胞的进一步分析表明,它们都是常规的B-2细胞,而B-1a(CD5+)B细胞仍然受到抑制。兔B细胞在功能上更类似于个体发育早期分化并自我更新的小鼠B-1细胞,而不是同样在发育早期产生但持续分化补充的B-2细胞。
分子水平上检测也证实了成年兔B细胞的生成有限。如果骨髓中正在进行B细胞生成,那么RNase保护试验,以种系VH基因作为探针,应该可以检测到重排后未分化的VDJ基因的mRNA。研究人员使用了源自种系VH基因的VH1 RNA探针,VH1是大多数兔VDJ基因重排中使用的VH基因,检测了从新生兔和成年兔骨髓中分离的mRNA。与预期结果一致,在幼兔骨髓中发现了保护VH1探针的mRNA,但成年兔骨髓中几乎不含能够保护探针的mRNA。这也就是说,成年兔骨髓中缺少未分化VDJ基因mRNA,即B淋巴细胞生成在成年兔中是有限的。
鉴定B细胞正在发育的另一种方法是寻找IgH基因座重排的证据。V、D和J基因重排使得与每个基因片段相关的重组信号序列(recombination signal sequence,RSS)重组形成带有信号连接产物的环状DNA。有研究证实,新生兔的骨髓细胞中存在大量此类环状DNA,但成年兔的骨髓中仅鉴定出少量。这些数据表明,成年兔骨髓中很少发生VD和DJ基因重排。成年兔脾脏、派尔集合淋巴结、肠系膜淋巴结、肠道或阑尾中同样没有持续重排的迹象。
如果成年兔B细胞的生成有限,则必须通过其他机制维持外周B细胞的数量。兔子似乎有不同的策略来维持不同抗体库所需的外周B细胞数量。研究者猜测兔B细胞是长寿的或自我更新的,或两者兼而有之。他们使用MHC基因座匹配的远缘兔建立了兔嵌合体。将成年脾细胞移植到MHC匹配但IgH同种异型不匹配的新生受体中后,观察到供体B细胞在受体中持续长达20个月。此外,研究人员使用成年兔骨髓作为供体细胞来源进行了移植实验,他们发现骨髓移植也能够在新生受体中成功,B细胞移植成功是因为骨髓中存在成熟的B细胞,而不是B细胞前体。这些实验表明,兔B细胞可能是长寿的或自我更新的。
兔B细胞的另一个有趣的点是它们都表达表面糖蛋白CD5。如前所述,小鼠CD5+B细胞(B-1a细胞)及其CD5–细胞(B-1b细胞)在表型和功能上均与B-2细胞不同。大多数B-1细胞是在胎儿和新生儿生命早期产生的,在成人骨髓中很少发现其前体。然而,B-1细胞确实在小鼠的一生中持续存在,并且基于其在新生小鼠和辐射小鼠中的扩增,它们确实是自我更新的。因此,如果兔B细胞在功能上与小鼠CD5+B细胞相似,那么它们也可能是自我更新的(有待直接检验)。
参考文献
[1]Knight KL, Winstead CR. B lymphocyte development in the rabbit. Int Rev Immunol. 1997;15(3-4):129-63. doi: 10.3109/08830189709068174.
[2]Jasper PJ, Zhai SK, Kalis SL, et al. B lymphocyte development in rabbit: progenitor B cells and waning of B lymphopoiesis. J Immunol. 2003 Dec 15;171(12):6372-80. doi: 10.4049/jimmunol.171.12.6372.
单克隆抗体是医学和生物学中最重要的生物工具之一,它彻底改变了一些疾病的诊断、治疗和靶向给药的方法。单B细胞抗体制备技术是单克隆抗体制备方法中最为直接但技术上最具挑战性的方法。该技术从单个B细胞直接“采样”免疫系统,从大量原代B细胞中检测和筛选特定的B细胞,扩增抗体基因,将基因克隆到表达载体中,并在哺乳动物细胞(如HEK293和CHO细胞)或细菌系统(大肠杆菌)中表达单克隆抗体。单B细胞抗体制备技术现已广泛用于生产人、小鼠、大鼠和兔单克隆抗体。本文以兔单克隆抗体为例,介绍使用单B细胞技术制备单克隆抗体的方法。
可用蛋白或与KLH偶联的多肽免疫新西兰白兔。多肽的序列应是目标抗原所特有的,并且与其他蛋白具有最小的同源性。每种免疫原至少免疫两只兔子。初次免疫皮下注射完全弗氏佐剂(CFA)乳化后的免疫原。加强免疫的免疫原与不完全弗氏佐剂(IFA)混合制备而成。通常在初次免疫前采集兔子血清样本作为对照,用于评估之后抗血清效价。每次加强免疫后1周采集血清样本,通过ELISA、免疫印迹或免疫组化试验测试多克隆抗体的水平及其功能。选择测定结果较好(强特异性信号和低背景信号)的兔子进行B细胞分选。
分选抗原特异性B细胞的方法很多。通过显微操作从组织中挑选标记过的B细胞和基于表面标记物的荧光激活细胞分选术是用于随机筛选B细胞的常用方法。流式细胞术也常用于分选抗原特异性B细胞。其他方法包括通过抗原包被的磁珠或微孔芯片阵列进行筛选,产生荧光焦点以分离抗原特异性浆细胞。
流式细胞术是分选抗原特异性B细胞最为有效的技术。这项技术已经成熟应用在人类B细胞的分选。人类B细胞表面上的多种标记物,可用于检测并分选免疫细胞的亚群。
使用流式细胞术从免疫动物中分选B细胞的方法与此相似。如利用流式细胞术分选源自小鼠脾脏的单个IgG+记忆B细胞:使用抗CD45R(B220)试剂富集小鼠B细胞,然后用抗CD19和抗IgG抗体进行标记,最后将荧光素标记抗原用于流式细胞术中特异性记忆B细胞的准确检测。
由于兔B细胞标志物较为缺乏,通过流式细胞术的鉴定和分选更具挑战性。在这种情况下,兔B细胞仅限于使用荧光标记抗原识别。同时,也可以使用反筛的方法,标记不需要的细胞,如T细胞和IgM+细胞的标记物,可以帮助分选到所需的IgG+细胞。
为了克服表面标记的局限性,有研究者采用了不同的策略。他们用抗IgG抗体和内质网(ER)特异性荧光染料对淋巴结细胞进行染色,如果这些细胞含有大量ER以产生抗体,则会被染色。他们将这种方法称为基于ER的抗原特异性浆细胞(ERIAA)鉴定,并将这种策略应用于鉴定人、小鼠、兔、大鼠和豚鼠的浆细胞。
扩增cDNA是B细胞分选后的下一步。如果培养B细胞,则将其裂解,从裂解物中提取总RNA和mRNA。如果未培养分选细胞,则将从单个细胞扩增cDNA。在逆转录聚合酶链反应(RT-PCR)过程中,通过使用随机六聚体引物、oligo-dT或基因特异性引物逆转录mRNA合成cDNA。通常,单个细胞在96孔板中分选,细胞裂解和cDNA合成在细胞分选的原始孔中进行。这样更容易处理大量样品。然而,在移液过程中,必须注意尽量减少相邻孔之间交叉污染的风险。
VH和VL的全长基因转录物通常在两轮PCR期间通过巢氏或半巢氏RT-PCR以第一链cDNA为模板扩增。正向引物可以是与相应的重链和轻链前导序列互补的序列或序列组合。反向引物可以是重链和轻链恒定区域内的序列或序列组合。例如,在兔IgG中,IgG只有一个亚类,重链恒定区只有一种基因。而兔IgG轻链70%以上为κ1,其余为κ2和λ。因此,在设计PCR引物时需要考虑这些细节。使用巢氏引物的第二轮扩增是为了增加特异性,并为随后的可变区克隆增加限制性位点。
克隆扩增可变区基因,表达抗体的方法有多种。一种策略是将PCR片段克隆到带有表达元件的质粒载体中,该表达元件包含前导序列和重链或轻链的恒定区。载体还需要具有转录和翻译所需的所有必要元素,如启动子序列和poly (A)序列。将载体转化到大肠杆菌中以制备足够的DNA用于转染。然后,将含有重链和轻链表达基因的质粒共转染到哺乳动物表达细胞中。重链和轻链由细胞表达,并组装为完整抗体结构释放到培养基中。
不依赖序列和连接的克隆方法(SLIC)是克隆编码可变区片段的一种不同方法。将携带重链或轻链恒定区序列的质粒,所有转录和翻译所需的元素都用限制酶消化。使用外切酶如T4 DNA聚合酶在该线性质粒和包含VH和VL的DNA片段中产生互补单链DNA突出末端。然后将它们在体外结合以产生HC和LC表达质粒。将质粒转染哺乳动物细胞,如HEK293进行抗体表达。
适当的培养时间后收获抗体上清液,确定产生抗体的数量和活性。可通过抗原结合ELISA试验确定抗体的特异性,也可通过免疫印迹进行检验。
在过去一两年中,单B细胞技术作为一种创新技术,已用于生产多种应用的单克隆抗体。与生产单克隆抗体的其他技术相比,单B细胞技术具有几个优点:
1. 与杂交瘤技术中低效的融合不同,单B细胞技术极为高效,并且保留了大部分抗体库。这有利于产生具有更高多样性的抗体。
2. VH和VL均从单个细胞中扩增。与展示方法不同,保持了抗体的天然同源配对和发育,保证获得抗体具有更高亲和力、稳定性和特异性。
3. 单B细胞技术会对VH和VL的DNA序列进行测序。因此,生产过程不依赖于细胞株,不存在污染和失去产生抗体的细胞的风险。
4. 不需要保存B细胞。
表1 常用抗体制备方法的优劣势对比
| 技术 | 优势 | 劣势 |
|---|---|---|
| 杂交瘤技术 | 抗体重轻链天然配对 |
1. 许多物种没有可用的融合伴侣细胞 2. 细胞融合效率低 3. 抗体制备所需时间久 |
| 展示技术 |
1. 无物种限制 2. 可通过控制筛选策略, 选择所需特异性的抗体 |
1. 抗体重轻链非天然配对 2. 建库较为困难 |
| B细胞永生化技术 | 抗体重轻链天然配对 |
1. 需要使用病毒将B细胞永生化 2. 细胞有污染的风险 |
| 单B细胞技术 |
1. 高效率 2. 抗体重轻链天然配对 3. 无需培养B细胞 4. 保留抗体多样性 |
技术门槛高 |
参考文献
Rashidian J, Lloyd J. Single B Cell Cloning and Production of Rabbit Monoclonal Antibodies. Methods Mol Biol. 2020;2070:423-441. doi: 10.1007/978-1-4939-9853-1_23.
自1975年,Milstein和Kohler成功融合出第一株鼠杂交瘤细胞后,单克隆抗体便开始进入大众视野,在医学检验、蛋白提纯、靶向治疗和显像技术中得以应用。1986年,第一款单克隆抗体药Orthoclone OKT3经FDA审批通过后上市,该药物靶向人CD3,用以防止肾脏移植后的宿主排斥问题。Orthoclone OKT3也成为了最早的第一代鼠源单抗药物。
经过47年的研究与发展,如今已有上百种单抗药物通过FDA审批上市,针对包括肿瘤、自免疫病、传染病等不同类型疾病。尽管仅过了人源化改造,绝大多数单抗药物仍来源于对鼠进行免疫后产生的鼠源抗体。鼠源单抗虽然制备技术相对成熟,发展历史相对悠久,但由于鼠本身脾脏较小,免疫系统产生的抗体多样性较差抗体种类较少的天然劣势,不少企业在开发抗体平台时会设法对鼠单抗进行改良;也有不少公司转而研发兔源单克隆抗体药以寻求突破。目前,已有两款兔源单抗药成功上市,分别是来自诺华(Novartis)的Beovu(Brolucizumab)和来自灵北(Lundbeck)的Vyepti(Eptinezumab)。
在详细介绍Beovu前,需要补充介绍一些兔单抗开发技术发展的背景知识。鼠源杂交瘤技术是利用鼠浆细胞与鼠源骨髓瘤细胞同源融合来获得鼠源杂交瘤的。然而,兔浆细胞与鼠源骨髓瘤细胞形成的兔-鼠异源杂交瘤细胞分泌抗体的效率较低,且异源杂交瘤并不稳定,极易退化。直到1997年,由朱伟民博士筛选出世界上第一株用于产生稳定、高质量的兔-兔单隆抗体的融合细胞株。该细胞株被命名为240E-W,由朱伟民博士和余国良博士建立的Epitomics公司掌握使用该细胞株制备兔单抗杂交瘤(RabMAb®)的专利。后来Epitomics公司经历了数次改组与收购,最终RabMAb®的专利落入了Abcam与诺华等公司手中。而借由兔单抗平台,诺华最终成功研发出Beovu并上市。

图1 兔抗药物发展史
Beovu(Brolucizumab)上市于2019年,是一种人类血管内皮生长因子(VEGF)抑制剂的scFv类兔单抗药,被FDA批准用于治疗湿性年龄相关性黄斑变性(AMD)。

图2 Beovu(Brolucizumab)
湿性年龄相关性黄斑变性,也称为渗出性或新生血管性年龄相关性黄斑变性,是由玻璃膜疣等引起的Bruch膜损害,能诱发脉络膜的毛细血管向外层长出新生血管,引起严重的视力丧失。血管内皮生长因子是一个家族,包括VEGF-A、VEGF-B等。VEGF-A可促进新生血管形成和使血管通透性增加,然而过多的VEGF-A则是湿性AMD产生的原因之一。Brolucizumab通过阻断VEGF-A与内皮细胞表面VEGF受体结合,减少新生血管病变,帮助缓解湿性AMD的症状,从而减缓视力下降。

图3 Brolucizumab阻断VEGF-A与VEGF受体结合
Beovu是一种单剂量瓶装的溶液(液体混合物),以注射方式给药。由于Brolucizumab为高度人源化兔单抗scFv,其半衰期较长,仅需在最初的三次药物治疗保持每月一次的频率,后续每8至12周给药一次。

图4 Beovu注射液
另一方面,Vyepti(Eptinezumab)作为唯一一款批准上市的全长兔单抗药物,最早由Alder BioPharmaceuticals(简称Alder)公司所研发。2019年,Alder被丹麦企业灵北制药(Lundbeck)以19.5亿美元的价格收购。2020年2月,Vyepti作为全球首款全长兔单抗药物上市,用于预防性治疗成人偏头痛。
偏头痛是一种可引起严重搏动性疼痛或搏动感的头痛,通常在头部的一侧。它常常伴随着恶心、呕吐,以及对光线和声音极度敏感。偏头痛发作可持续数小时至数天,疼痛程度可能非常严重,以至于影响日常生活。降钙素基因相关肽(Calcitonin gene related peptide, CGRP)是人类用分子生物学方法发现的第一个活性多肽。CGRP的释放会引起局部血管扩张,尽管具体的机理尚不明确,但它仍被认为在偏头痛中起着关键作用。

图5 CGRP与偏头痛
自2018年以来,美国FDA已经批准了6种可以阻断CGRP或其受体的药物,Vyepti便是其中之一。这些药物副作用相对较小,而且对于许多接受其他药物治疗无应答的人群有效。Vyepti通过与CGRP结合,从而阻断CGRP与其受体的结合过程来缓解偏头痛的症状。由于CGRP仅由37个氨基酸组成,相对较小,Eptinezumab作为兔单抗,对小分子和半抗原也会产生更明显的免疫反应。

图6 Vyepti作用机制
目前针对CGRP已有四款抗体药物上市,除Vyepti外,分别为安进/诺华Aimovig、梯瓦Ajovy、礼来Emgality。相比其他三种鼠源抗体药物,Vyepti是唯一一种静脉给药剂型。用药方面,Vyepti每3个月一次静脉输注,Aimovig和Emgality均为每月一次皮下注射,Ajovy则可每月一次或每3个月一次皮下注射。Vyepti给药周期明显较长,更能减少患者依从性问题。
相比鼠源单抗,兔单抗具备特异性更高,亲和力更好,识别表位更精细等优势。同时,越来越多的小鼠疾病模型也被投入到临床试验中。由于鼠源单克隆抗体不识别鼠源蛋白,更多的兔源单抗替代药也逐渐进入视野。种种原因下,更多生物药企开始从事针对更多不同靶点的兔单抗药物研发,如国内奥赛康药业针对胰腺癌Claudin 18.2的ASKB589,荃信生物针对银屑病IL-17A的QX002N等以及国外Apexigen针对CD40的APX005M等。随着更多的兔单抗药物进入临床阶段,如何得到特异性针对新靶点并能成功成药的兔单克隆抗体分子,也将成为诸多生物药企业的一大攻克目标。

图7 部分进入临床阶段兔单抗药物名单
与此同时,在兔单抗药物先导分子发现阶段,新时代的方法与技术也在不断更新,从而摆脱兔抗药研发受RabMAb®专利的影响。第二代的抗体展示技术可以避免兔单抗免疫耐受的问题,并可针对κ型抗体,也可用于λ型的高亲和力抗体筛选。最新的单B细胞技术通过直接对单个B细胞的筛选,可在不到两个月的时间,筛选出大量的候选兔单抗分子。新平台与旧方法的碰撞,也提供给更多兔单抗研发药企更多选择。

图8 SingleB兔单抗制备流程
另一方面,虽然兔抗的同源性相较人的同源性比鼠相对较高(约60%~76%),并且最近新开发的Mutational Lineage-Guided(MLG)人源化技术也使得RabMAb人源化更加容易,但由于兔单抗重链可变区含有额外的二硫键,兔单抗的人源化仍然是兔单抗分子成药的一大挑战。新平台的开发或者转基因兔的培育或许会成为未来探索的重点。
兔单抗的诞生是曲折的,相比鼠单抗,无论是发展历史还是上市药数量,都相较式微。但兔单抗的高亲和,高特异等优势为生物医学研究和抗体药物的发展提供了大的空间。兔单抗药研发的道路依然十分漫长,但基于新的平台,攻克新的困难,也必然会指向兔单抗药光明的未来。
参考文献
[1]Justus Weber, Haiyong Peng, Christoph Rader. From rabbit antibody repertoires to rabbit monoclonal antibodies. Experimental & Molecular Medicine 2017 (49)
[2]Thomas FT, Griesedieck C, Thomas J, Carver M, Whitley T, Warren R et al. Differential effects of horse ATG and rabbit ATG on T cell and T cell subset levels measured by monoclonal antibodies. Transplant Proc 1984; 16:1561-1563.
[3]Zhu L, Fang Y, Liu Z, Wang P, Wang Y, Xu H. Rabbit anti-human leukocyte polyclonal antibody inhibits xenogeneic cell-mediated immune responses. Transplant Proc 2008; 40: 2760-2763.
[4]FDA. Available at http://www.accessdata.fda.gov/scripts/cdrh/devicesatfda/index.cfm (using search term ‘rabbit polyclonal antibodies’, accessed on 3 October 2016).
[5]Reichert JM. Antibodies to watch in 2016. MAbs 2016; 8: 197-204.
[6]Weiner GJ. Building better monoclonal antibody-based therapeutics. Nat Rev Cancer 2015; 15: 361-370.
伴随诊断兔单抗:新时代、新选择
非特异性:治疗性抗体开发中的难题
免疫球蛋白分子的结构和稳定性取决于于免疫宿主动物的类别和亚类。与啮齿动物相比,兔子对小分子、精细表位也可产生有效的免疫反应,抗体产量高,获得高亲和力抗体的机率更大。兔抗体分子的稳定与轻链的多样化是密不可分的。兔子只有一个γ免疫球蛋白(IgG)同型,其重链可变区的VH1种系使用率约为80-90%。但兔子轻链是多样化的,除了λ链,还有b4、b5、b6和b9四个κ异型。兔子κ链Cys80和Cys170之间存在一个二硫键,将κ链的可变区(Vκ)与恒定区(Cκ)连接在一起(图1a),这个域间二硫键是兔抗结构特有的。
有研究人员使用三个κ-b4轻链同种异型的单克隆IgG(A3、A4、G11),通过定点突变,用Ala取代Cys80和Cys170产生IgG-AA。通过差示扫描量热法(DSC)和等温滴定量热法(ITC)测定了IgG-AA与IgG的热稳定性和结合活性参数,分析了这种Vκ-Cκ域间二硫键在IgG稳定性和抗原相互作用中的作用。

图1 兔κ链可变区和恒定区之间的二硫键
注:(a)兔子Fab结构,重链(H)和kappa链(κ)的可变区(V)和恒定区(C)分别标记为VH(绿色)、CH1(灰色)、Vκ(浅绿色)和Cκ(洋红色)。二硫键以黄色显示。Vκ和Cκ结构区之间独特的二硫键显示在右框中。(b)三个IgG κ链二硫键所在位置的氨基序列。两个保守的半胱氨酸用黄色标出。(c)IgG和IgG-AA示意图。
兔子单克隆抗体κ链中的二硫键有助于提高抗体的热稳定性,二硫键的存在不影响抗体结构域的折叠及结构域间的接触,仅从动力学和熵变两方面影响IgG的热稳定性。实验结果显示,二硫键对整体结构没有显著影响,所有三种抗体(IgG和IgG-AA)尺寸排阻层析(SEC)检测的洗脱曲线几乎相同。三种同种异型抗体(包括IgG和IgG-AA)熔解曲线计算的表观量热展开焓(ΔHm)几乎相同。热变性重新扫描显示,IgG-AA第一次变性时熔解温度(Tm)的降低幅度大于其他峰。G11组扫描结果显示,扫描速率越小,变性开始越早,Tm也越低,且IgG-AA组的变化更大。表明兔抗体二硫键的热稳定不是热焓变化的结果,这种热稳定性主要取决于动力学稳定性,抗体热变性时域间二硫键会成为能量屏障,干扰热变性的初始步骤。

图2 A4、G11和A3的IgG和IgG-AA DSC热分析图
注:IgG和IgG-AA的结果分别用虚线和实线表示;(a)A4 IgG浓度为17.9 μM,A4 IgG-AA为14.8 μM;(b)G11 IgG为24.5 μM、G11 IgG-AA为25.3 μM;(c)A3 IgG为16.7 μM、A3 IgG-AA为14.1 μM。
表1 熔解中间温度(Tm)和变性焓(ΔHm)

注:通过扫描1.0°C/min的DSC测量所得。
通过ITC分析兔IgG和IgG-AA与抗原的结合活性,获得的热力学参数示如表2所示。3种同种异型IgG和IgG-AA与抗原的解离常数差异非常微小,相互作用的焓变(ΔH)或熵变差异也非常小,表明二硫键对抗体结构无显著影响,对抗体-抗原相互作用的非共价键形成(ΔH°)、水合作用和灵活性的变化(−TΔS°)影响也非常小(图3和表2)。

图3 A4 IgG(左)和IgG-AA(右)与ITC分析的抗原相互作用
表2 热力学参数

注:通过ITC测量热力学参数,25°C时的参数显示为三次测量的平均值±s.d.。
兔抗体中连接Vκ和Cκ的二硫键不影响抗原抗体相互作用,仅在IgG不可逆热变性过程的动力学稳定中起作用。类似的稳定二硫键可以有效地合并到具有高拓扑相似性的其他抗体(亚)类中。嵌合体和人源化兔抗体的相关研究发现,这种二硫键确实可以提高抗体的热稳定性。兔抗体中κ轻链域间二硫键的发现,有助于指导耐高温稳定抗体的工程设计,以供临床开发。
参考文献
Raiji Kawade, Hiroki Akiba, Kevin Entzminger, et al. Roles of the disulfide bond between the variable and the constant domains of rabbit immunoglobulin kappa chains in thermal stability and affinity[J]. Protein Engineering, Design and Selection, 2018, 31:243–247.






南京德泰生物工程有限公司 Nanjing Detai Bioengineering Co.,Ltd. ©2026 All Rights Reserved
 苏公网安备32011202001300
苏公网安备32011202001300