
抗体是体液免疫的最主要成分,是免疫系统在抗原刺激下,由B淋巴细胞或记忆B细胞增殖分化形成的浆细胞产生的、可与相应抗原发生特异性结合的免疫球蛋白(Immunoglobulin, Ig)分子。作为具有生物学功能的大分子蛋白质,抗体既可以通过其抗原结合部位识别并结合相应的抗原,同时也具有抗原性,可以刺激机体产生相应的抗体。
由于抗体产生的遗传基础不同,因而不同抗体分子的抗原性存在差异。利用血清学方法可以测定和分析不同抗体分子的抗原性并将其进行分类,称之为抗体的血清型。抗体的血清型可以分为同种型(Isotype)、同种异型(Allotype)和独特型(Idiotype, Id)。
 |
同种型,指同一种属所有个体抗体分子共有的抗原特异性标志,其表位存在抗体的恒定区。 |
 |
同种异型,指同一种属不同个体间抗体分子所具有的不同抗原特异性标志,其表位广泛存在于抗体的恒定区。 |
 |
独特型,指每个抗体分子所特有的、存在于可变区的抗原特异性标志。独特位可位于抗体分子V区的互补决定区(CDR),也可位于抗体分子V区的骨架区(FR)。 |
独特型由若干表位组成,称为独特位(idiotope),它可刺激机体产生相应的抗体,即抗独特型抗体(anti-idiotypic antibody, AId)。独特型和抗独特型抗体可以形成复杂的网络,在免疫调节中占有重要地位。
根据抗独特型抗体与独特型反应的特点,可将抗独特型抗体分为Ab2α、Ab2β、Ab2γ、Ab2δ四类。
| Ab2α | 由位于Id骨架区的独特位诱导产生,与Id的结合不影响与抗原的结合,属于抗原非抑制性的抗独特型抗体。 |
| Ab2β | 由位于Id抗原结合位的独特位诱导产生,这类抗独特型抗体在三维结构上与抗原类似,Ab2β和Id的结合可以完全阻断抗原与Id的结合,这类抗独特型抗体在实际应用上具有重要意义。 |
| Ab2γ | 由靠近抗原结合位的独特位诱导产生,与Id的结合可以抑制抗原与Id的结合。与Ab2β不同的是,Ab2γ在三维结构上与Id原始抗原的结构并不相似。Ab2γ在与Id结合后,可导致Id可变区三维结构改变,干扰抗原与Id的结合。 |
| Ab2δ | 既能识别Id骨架结构上的独特位,同时也能识别抗原上具有交叉反应结构的表位,是一类比较少见的抗体。 |
根据结合模式的不同,可将抗独特型抗体分为抗原阻断型、抗原非阻断型和药物靶标复合物型。
互补位特异性
阻断抗原抗体结合
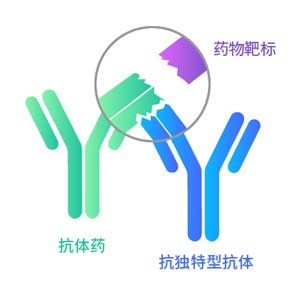
互补位非特异性
不阻断抗原抗体结合
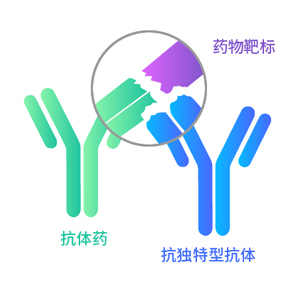
特异性结合药物靶点复合物
不阻断抗原抗体结合
根据免疫网络学说,作为“内影像”的抗独特型抗体在结构和功能上能模拟相应抗原,在一定程度上替代原始抗原,因此可以将抗独特型抗体作为抗原的替代物用于免疫学研究及基础和临床医学研究,而且在传染性疾病的防治机理研究和肿瘤治疗等应用研究中也取得了重大进展。
1.寻找病毒受体
病毒受体指位于宿主细胞表面能够被病毒吸附蛋白识别并与之结合,从而引起病毒感染的分子复合物,大多数属于蛋白质。病毒受体是公认的引发病毒感染宿主细胞的主要决定因素,也是影响病毒宿主特异性和组织亲嗜性的决定因素之一。根据Jerne的免疫网络学说,作为抗原“内影像”的抗独特型抗体在结构和功能上能模拟相应的外来抗原,因此能够用于模拟病毒寻找细胞受体蛋白。通常选择病毒的中和抗体制备抗独特型抗体,所获得的抗独特型抗体能很好地模拟病毒抗原。
2.制备受体拮抗剂
受体拮抗剂指能与受体结合,但不具备内在活性的一类物质,也指与受体有较强的亲和力而无内在活性的药物。例如传统的生长激素受体拮抗剂主要分为两类,生长激素类似物和抗生长激素受体抗体。抗独特型抗体可以代表一种制备拮抗剂的新策略并应用于其他细胞因子和生长因子拮抗剂的研究。
3.疫苗研究
抗独特型抗体Ab2β作为抗原的模拟物,可代替病原体诱导机体产生抗病原体的特异性免疫应答,即所谓抗独特型疫苗。抗独特型疫苗是免疫调节网络学说发展到新阶段的产物,无论抗原是细菌、病毒还是寄生虫,或是蛋白质还是多糖,都可用其“内影像”抗体来诱导机体的特异性免疫应答,从而产生保护性免疫应答。目前,已有许多用抗独特型抗体制备的实验室疫苗接种动物后能使免疫动物抵抗病原体的感染。
除病原防治领域外,抗独特型肿瘤疫苗的研究也逐渐成为研究热点。无论是最初的鼠源性Ab2β,还是Ab2β的Scfv疫苗以及与DC、细胞因子等联合制成的疫苗,抗独特型抗体的功能在不断得到改进和完善,在大多数动物试验研究中都能诱导出抗肿瘤的特异性细胞免疫和体液免疫,在临床试验中也取得了较好的疗效。
4.诊断试剂
抗原是临床免疫诊断中最为关键的试剂之一,但由于某些天然抗原的自然含量极少,或者纯化抗原极其困难,或者抗原本身具有危险性等原因,难以获得必要的天然抗原。用抗独特型抗体替代这些抗原作为诊断试剂,不仅可以克服这些困难,而且具有重复性强、稳定性好、可标准化等优点。
5.自身免疫病研究
自身免疫病是机体产生对自身成份的免疫应答并造成病理损害而引起的一系列疾病,常与自身隐蔽抗原的释放、自身反应的T细胞克隆激活以及由于具有与自身交叉反应抗原的病原体感染有关。随着免疫网络学说和抗独特型抗体研究的发展,进一步提示了独特型网络维护机体免疫系统自身稳定的一个重要功能,如果调节失常,就可能导致自身免疫病的发生。
6.药代动力学检测
抗体药是生物技术药物的重要组成部分,其药代动力学(PK)检测是生物技术药物开发过程中的重要挑战之一。PK检测通过检测抗体药在动物或人体内的血药浓度-时间曲线、组织分布、代谢转化排泄等,对药物的临床安全性及有效性进行评估,确定药物初始使用剂量,指导临床用药。抗独特型抗体是抗体药PK分析的关键工具试剂,可以用于检测血液中抗体药的含量,包括游离型、结合型及总量。抗原阻断型抗独特型抗体可以检测游离型抗体药,抗原非阻断型抗独特型抗体可以检测抗体药总量,而药物靶标复合型可以检测检测结合型抗体药。
7.免疫原性分析
药物的免疫原性,通常是指其诱发抗药抗体(Anti-drug antibody, ADA)反应的能力。抗体药作为外源性蛋白,不可避免地会激活机体自身地免疫反应,诱导产生抗药抗体,从而影响抗体药的安全性和有效性。抗体药人源化可以减少非人源成分带来的各种风险,但是仍会产生一定的抗药抗体。
免疫原性分析是生物技术药物临床研发中的重要部分,通常是通过对药物引起的抗药抗体的检测和确认来评估药物的免疫原性。有效评估抗体药物的免疫原性,开发可靠的评估检测方法尤为重要。免疫原性筛选通常使用操作方便快捷、能够高通量测定的ELISA方法。其中桥式ELISA法最为常用,该方法是利用包被在酶标板上的抗体药物捕获血清样品中的ADA,然后加入生物素标记的抗体药物,使其与ADA结合。接者用链霉亲和素-辣根过氧化物酶偶联试剂作为检测试剂,加入酶反应底物显色后,再利用酶标仪进行检测。此法的优点是能检测各类抗体的亚型,可高通量检测。在分析患者样本中是否存在抗药抗体时,常使用多克隆抗独特型抗体作为阳性对照。
细胞因子释放相关综合征(CRRS)常见于T细胞抑制剂(TCE)的临床试验中,通常由T细胞过度活化引发,进而影响药物的理想分布和长期疗效。细胞因子的产生是T细胞活化的标志吗?或者,在临床环境中,高水平的细胞因子分泌是TCE疗效的必要条件吗?
这两个问题的答案都是否定的。研究揭示,激活细胞毒性T淋巴细胞(CTL)的生物功能是由CTL与靶点之间的分子动力学决定的,存在一个阈值实现完全激活。
开创性的研究结果表明,T细胞杀伤并不需要形成稳定成熟的免疫突触,并为下一代TCE的开发提供了将细胞毒性与毒性触发的T细胞刺激解耦的基本原理,在下一代TCE中,CD3结合亲和力被微调(通常是减弱),以达到一个信号的”甜蜜点”,从而在不产生过多细胞因子释放和/或T细胞衰竭的情况下推动抗肿瘤细胞溶解活性。

迄今为止,许多新型TCE都是基于这一概念进行构建的,所产生的数据都支持“解耦”理论,以下例子候选TCE药物一一进行具体阐述。
这款CD19×CD3双特异性抗体是专为治疗B-NHL而设计的。虽然其CD3靶向臂的亲和力较低,降低了体外杀伤力,但却能达到相同水平的最大杀伤,而且刺激细胞因子释放的量明显较少。令人惊奇的是,仅在低剂量下就能在体内长期杀死肿瘤细胞。CD3低亲和力导致的有利生物分布,或诱导T细胞衰竭或活化诱导细胞死亡(AICD)的倾向较少,可能是其改善体内功能的原因。

这种新型TCE代表了一种新型疗法,它能诱导T细胞介导的强大肿瘤细胞毒性,与高水平的细胞因子释放无关,因此是治疗B-NHL的理想候选药物。
这款与CD3结合亲和力较低的HER2/CD3 TCE可降低全身细胞因子的释放,及对正常组织的脱靶毒性,证明了低亲和力可提高动物模型的总体耐受性,而T细胞结合亲和力对体外和体内抗肿瘤活性的影响有但非常有限。这些数据表明,对CD3和肿瘤靶点的亲和力进行微调是实现CD3 TCEs最大治疗指数的一种有前途的策略。

这款经过CD3亲和力优化的TCE显示出了杀死癌细胞和引发T细胞增殖的能力,但细胞因子的释放量有所减少,而且有可能在多发性骨髓瘤患者身上激发显著的临床活性。

这款用于治疗卵巢癌的CD3亲和力定制MUC1×CD3 TCE表明,细胞因子的释放曲线有所减弱,生物分布良好,但其杀瘤效力依然存在,这表明REGN4018具有强大的抗肿瘤活性和良好的耐受性,目前正在进行临床评估。

这家生物技术公司正在通过微调CD3结合亲和力和疾病靶标结合分子,确定抗肿瘤效力和毒性(主要是CRRS)平衡的TCE。其概念验证TCE已显示出高效力和低细胞因子释放。
德泰生物拥有尖端的单B抗体发现平台,致力于发现靶向细胞溶解效应细胞刺激受体的高质量定制激动抗体(agonistic antibodies),以构建下一代多特异性抗体。
参考文献
[1]Faroudi, Mustapha, et al. “Lytic versus stimulatory synapse in cytotoxic T lymphocyte/target cell interaction: manifestation of a dual activation threshold.” Proceedings of the National Academy of Sciences 100.24 (2003): 14145-14150.
[2]Riquelme, Erick, et al. “The duration of TCR/pMHC interactions regulates CTL effector function and tumor‐killing capacity.” European journal of immunology 39.8 (2009): 2259-2269.
[3]Malik-Chaudhry, Harbani K., et al. “TNB-486 induces potent tumor cell cytotoxicity coupled with low cytokine release in preclinical models of B-NHL.” MAbs. Vol. 13. No. 1. Taylor & Francis, 2021.
[4]Staflin, Karin, et al. “Target arm affinities determine preclinical efficacy and safety of anti-HER2/CD3 bispecific antibody.” JCI insight 5.7 (2020).
[5]Zuch de Zafra, Christina L., et al. “Targeting multiple myeloma with AMG 424, a novel anti-CD38/CD3 bispecific T-cell–recruiting antibody optimized for cytotoxicity and cytokine release.” Clinical Cancer Research 25.13 (2019): 3921-3933.
[6]Crawford, Alison, et al. “A Mucin 16 bispecific T cell–engaging antibody for the treatment of ovarian cancer.” Science translational medicine 11.497 (2019): eaau7534.
目前,治疗性抗体在治疗癌症、肿瘤、自身免疫性疾病、慢性炎症、感染性疾病等领域取得了重大进展,抗体工程的发展进一步推动新型抗体的产生。截至2022年年底已上市的治疗性抗体药物及其衍生物超过100种。基于实用性和功能上的考虑,现在临床上使用的治疗性抗体大多为IgG类型。
在开发治疗性抗体时,IgG亚型的选择是基于其体外活性可以推断体内活性的假设。亚型是决定治疗性抗体的体内生物学活性的关键因素之一。除亚型外,IgG-Fc糖型、抗原决定簇密度、细胞Fc受体多态性等多个参数也共同影响着治疗性抗体体内功能结果,导致准确预测治疗性抗体的体内活性仍有困难。因此在抗体药物设计中应把亚型的结构特征与目的效应功能的关联、体外IgG-Fc和效应配体之间相互作用的分子机制、亚基的生物学活性、理化性质等纳入综合考虑,以确保最大的疗效和安全性。
虽然IgG各亚型在体外实验评估中呈现特定的效应功能,然而在不同疾病中各亚型体内的作用方式可能有所不同。因此,在治疗性抗体的开发中为达到期望的治疗效果并且避免不良反应,选择适宜的抗体亚型就显得格外重要。
IgG可根据其在血清中的分布丰度进一步分为IgG1(60%)、IgG2(25%)、IgG3(10%)、IgG4(5%)4种亚型,它们的恒定区序列高度同源。4种IgG亚型其恒定区的CH2区不同、铰链区的核心CXXC序列组成不同,重链和轻链之间的二硫键连接方式也不同。

图1 IgG亚型及其异构体示意图
(A)不同的IgG亚型重链和轻链结构、铰链的长度以及连接两条重链的二硫键的数量。
(B)显示了IgG2的B/B形式(重链间只有两个二硫键,并存在重轻链间的替代连接)以及中间A/B形式。
(C)IgG4的异构体导致半分子交换。
表1 IgG各亚型的结构差异
| IgG1 | IgG2 | IgG3 | IgG4 | |
|---|---|---|---|---|
| 分子质量(kDa) | 146 | 146 | 170 | 146 |
| 活性形式 | 二价单体 | 四价二聚体 | 二价单体 | 单价半抗体 |
| 同型 | 4 | 1 | 13 | 0 |
| 铰链区氨基酸个数 | 15 | 12 | 62a | 12 |
| 铰链区链内二硫键的个数 | 2 | 4b | 11a | 2 |
a:Depends on allotype;b:A/A isomer.
人类Fcγ受体家族包括六个成员,它们在细胞分布、与Fc的结合亲和力以及产生的生物活性方面有所不同。功能上最相关的FcγR是FcγRI,FcγRIIA,FcγRIIB和FcγRIIIA。FcγRI是一种对IgG具有高亲和力的激活受体,可以被单体IgG激活。除了FcγRI外,所有其他FcγR只有在抗体与特定抗原结合时,其Fc区才能激活下游效应功能。FcγRIIB是唯一的抑制性FcγR,在许多免疫细胞上表达,包括B细胞,单核细胞,巨噬细胞,DC和肥大细胞。当IgG与抗原结合时,FcγRIIB通过胞内免疫受体酪氨酸抑制基序(ITIM)转导抑制信号,抑制ADCC和ADCP。最典型的激活FcγR是FcγRIIA和FcγRIIIA。FcγRIIA主要在单核细胞和巨噬细胞上表达,而FcγRIIIA主要在NK细胞上表达。两种激活的FcγR都有两种等位基因变体。
与结构相适应,4种IgG亚型与抗原结合、免疫复合物的形成、补体激活、触发效应细胞、半衰期和胎盘转运特性均具有特异性。表2总结了IgG亚型功能特征的差异。
在4个IgG亚型中,IgG3与大多种类的FcγR的亲和力结合度最高,但由于其长铰链区和多态性,这两者都增加了稳定性和免疫原性的风险,未能成为治疗性抗体的常规选择。在其余亚型中,IgG1对所有FcγR表现出最高的亲和力,并且可以有效激活ADCC和ADCP反应。IgG4仅对FcγRI具有高亲和力,对其他受体的亲和力较弱,其Fc介导的效应功能较弱。IgG2对FcγRIIAH131形式具有高亲和力,但对FcγRI和其他FcγR没有可测量的或微弱的亲和力。
表2 IgG各亚型的功能差异
| IgG1 | IgG2 | IgG3 | IgG4 | |||||
|---|---|---|---|---|---|---|---|---|
| 体内生物学活性 | 蛋白质抗原 | 糖类抗原 | 蛋白质抗原 | 响应慢性刺激、抗炎活性 | ||||
| 半衰期(天) | 21 | 21 | 7~21a | 21 | ||||
| 胎盘转运 | ++++ | ++ | ++/++++a | +++ | ||||
| 与下列物质的抗体反应 | ||||||||
| 蛋白质 | ++ | +/- | ++ | ++e | ||||
| 多糖 | + | +++ | +/- | +/- | ||||
| 过敏源 | + | (-) | (-) | ++ | ||||
| 补体激活 | ||||||||
| 结合C1q | ++ | + | +++ | – | ||||
| Fc受体 | ||||||||
| FcgRI | +++ | 65d | – | – | +++ | 61 | ++ | 34 |
| FcγRIIaH131 | +++ | 5.2 | ++ | 0.45 | ++++ | 0.89 | ++ | 0.17 |
| FcγRIIaR131 | +++ | 3.5 | + | 0.10 | ++++ | 0.91 | ++ | 0.21 |
| FcγRIIb/c | + | 0.12 | – | 0.02 | ++ | 0.17 | + | 0.20 |
| FcγRIIIaF158 | ++ | 1.2 | – | 0.03 | ++++ | 7.7 | – | 0.20 |
| FcγRIIIaV158 | +++ | 2.0 | + | 0.07 | ++++ | 9.8 | ++ | 0.25 |
| FcγRIIIb | +++ | 0.2 | – | – | ++++ | 1.1 | – | – |
| FcRn (at pH < 6.5) | +++ | +++ | ++/+++a | +++ | ||||
a:Depends on allotype;b:A/A isomer;c:Multivalent binding to transfected cells;
d:Association constant (×106 M-1) for monovalent binding; e:After repeated encounters with protein antigens, often allergens
多达12个在肿瘤细胞上表达的靶标已经过临床验证。除靶向EGFR的西妥昔单抗和帕尼单抗分别选择IgG1和IgG2亚型外,所有其他靶标均选择了IgG1亚型。Her2和EGFR属于酪氨酸激酶受体家族,靶向抗体可能通过阻断生长信号发挥抗肿瘤活性,除此之外IgG1抗体诱导的针对受体酪氨酸激酶(RTK)的ADCC和/或ADCP也起着重要的治疗作用。因此,在开发靶向肿瘤细胞上靶标的治疗性抗体时,主要考虑IgG1,它对激活FcγR具有最佳的结合亲和力,可以引起针对肿瘤细胞的强ADCC和/或ADCP活性。此外,当ADCP被激活时,效应细胞(巨噬细胞或树突状细胞)可能会将处理后的肿瘤抗原呈递给T细胞,从而进一步引发持久的肿瘤特异性适应性免疫。
一些肿瘤细胞靶标是免疫检查点(ICP)的配体,例如PD-L1和CD47,它们分别是PD-1和SIRPα的配体。PD-1及其配体PD-L1通过逃避肿瘤中和免疫监视在肿瘤进展和存活中起至关重要的作用。PD-L1在肿瘤细胞和抗原呈递细胞中表达,与T细胞PD-1的结合导致肿瘤中的T细胞功能障碍、衰竭、中和和IL-10产生。因此,阻断PD-1或PD-L1识别过程可以阻断抑制信号传导,从而重新激活效应T细胞杀死肿瘤细胞。简单地阻断PD-L1可以诱导显著的治疗效果,但以IgG1形式保留Fc效应器功能将进一步提高治疗效果。据报道,阿维鲁单抗是一种IgG1亚型抗PD-L1抑制剂,可触发NK细胞介导的细胞毒性和细胞因子产生以对抗三阴性乳腺癌细胞。
ADCC的诱导取决于肿瘤细胞上靶标表达的丰度,每个细胞的最低需求为105~106。与肿瘤组织相比,正常组织通常表达水平要低得多。丰度小于105的细胞诱发可能小,因此不会受到ADCC的损害。
靶向免疫细胞靶标的IgG亚型选择更为复杂。由于细胞表达免疫抑制受体的分布不同,如PD-1、TIM-3和LAG-3主要在CD8 T淋巴细胞(免疫效应细胞)中表达,而CTLA-4主要在Treg细胞(免疫抑制细胞)中表达,在开发针对免疫抑制受体(ICP)的治疗性抗体时,应仔细考虑IgG亚型的影响。目前批准的所有PD-1抗体都是IgG4亚型形式,可见IgG4是PD-1抗体的首选形式。这是因为IgG4仅对FcγRI具有高亲和力,但对所有其他FcγR的亲和力较弱,不具有可检测的Fc介导的效应功能,这意味着CD8 T淋巴细胞不会被相应消除。
针对Tregs或其他免疫抑制细胞表达的ICP可以选择IgG1亚型抗体,这些细胞可以通过ADCC/ADCP被抗体清除。靶向CTLA-4的伊匹木单抗就是一种主要通过ADCC/ADCP消除TME中的Tregs细胞来发挥治疗效果的IgG1亚型抗体。而靶向CTLA-4的IgG2亚型替西木单抗自2006年以来一直在进行广泛的临床试验。由于IgG2仅对FcγRIIaH131具有高亲和力,FcγRIIaH131主要在巨噬细胞中表达,在没有CD47-SIRPα阻断剂的情况下不会完全激活,因此替西木单抗将无法从TME中消除Tregs细胞。
免疫刺激受体大多数属于肿瘤坏死因子受体(TNFR)超家族(如4-1BB,CD40,OX40,CD27,GITR),经激动抗体结合后可刺激免疫。针对ICP受体的抗体单纯阻断靶点可激活T细胞,而通过抗体激活TNFR超家族成员需要抗体与FcγR交联。这表明FcγRIIB是靶向TNFR超家族成员免疫刺激受体的抗体所需要的。此外,CH1结构域中的铰链区也很重要,是抗体激动功能所必需的。给定IgG的铰链越硬,免疫刺激受体在细胞膜上的聚集越稳定,抗肿瘤功效越好。
总的来说,在开发靶向肿瘤抗原的抗体药物时,应优先考虑IgG1亚型,特别是当Fc介导的效应功能是肿瘤治疗的主要作用方式时。在选择免疫细胞中靶标的IgG亚型时,应仔细考虑靶标的性质。如果ICP在免疫效应细胞上表达,则应优先考虑IgG4。如果在Treg或其他免疫抑制细胞上表达,则应选择IgG1。对于具有免疫刺激功能的靶标,特别是TNFR超家族成员的靶标,除了表位筛选外,还应设计增强FcγRIIB参与。
参考文献
[1]Vidarsson G, Dekkers G, Rispens T. IgG subclasses and allotypes:from structure to effector functions. Front Immunol, 2014, 5 (520) : 1–17.
[2]吕若芸,陈忱,魏敬双. 治疗性抗体药物开发中IgG亚型选择[J]. 中国生物工程杂志, 2016, 36(7): 104-111.
[3]Yu J, Song Y, Tian W. How to select IgG subclasses in developing anti-tumor therapeutic antibodies. J Hematol Oncol, 2020, 13: 45.
单克降抗体技术是现代生命科学研究的重要工具,自1975年杂交瘤单克隆抗体技术的出现,单克隆抗体迅速在生命科学研究和临床检测诊断中得到了广泛的应用,为许多领域的发展做出了不可估量的贡献。目前单克降抗体制备技术有杂交瘤技术、噬菌体展示技术以及单个B细胞抗体开发技术等。
单个B细胞抗体开发技术是近年来新兴的单抗制备技术。它是将单细胞分离鉴定技术结合多种PCR技术形成的一种单克隆抗体体外筛选及表达体系。单个B细胞抗体开发技术利用每个B细胞只产生一种特异性抗体的特性,使用流式细胞术、微流控技术等方法对免疫动物的B细胞进行抗原特异性单细胞分选,通过PCR扩增及一代、二代测序获得抗体基因序列,并在哺乳动物细胞中进行表达。
德泰生物基于单个B细胞抗体开发技术,自主研发了SingleB®单B细胞快速单抗发现平台。平台支持小鼠、大鼠、兔、羊驼、人源化小鼠、犬、羊、鸡等多物种抗体发现,通过高通量筛选获得抗原特异性记忆B细胞或浆细胞,单细胞PCR扩增及Sanger测序获取抗体基因序列,并进行重组表达与验证。平台可将抗体发现过程中的binding/blocking功能筛选前置,更快锁定阳性克隆,从动物免疫到获得单抗,快至29天,节省研发时间和费用。

杂交瘤技术是一种将B淋巴细胞与骨髓瘤细胞融合生产单克隆抗体的传统方法。该技术通过将动物脾细胞与骨髓瘤细胞融合,产生永生化杂交瘤细胞系,然后筛选其上清液中的抗原特异性克隆,并进一步循环亚克隆以产生严格的单克隆抗体。
利用杂交瘤制备单克隆抗体通常操作流程为:抗原制备、用抗原免疫动物、取免疫动物的脾脏细胞与骨髓瘤细胞融合成杂交瘤细胞、对杂交瘤细胞进行筛选及克隆化、利用筛选得到的杂交瘤细胞进行单克隆抗体的大量生产。了解杂交瘤制备单克隆抗体的详细流程。

利用展示平台实现文库系统表达和筛选目标分子是单抗开发的重要手段。以噬菌体展示技术为例,其原理是将一段外源基因插入到噬菌体外壳蛋白结构基因的适当位置,在阅读框正常且不影响外壳蛋白正常功能的情况下,外源基因会随着外壳蛋白的表达而表达,从而使多肽或蛋白以融合蛋白的形式展现在噬菌体表面。动物毒性抗原、自身抗原、免疫耐受原和弱免疫原性抗原,由于无法产生有效免疫,可采用展示技术获得的天然抗体库、合成或半合抗体库进行筛选。使用展示技术获得抗体通常为scFv(噬菌体表面展示制备scFv流程)、Fab形式而非全长抗体,且重轻链非天然配对,故亲和力和特异性较其他方法略有局限。

| 技术 | 优势 | 劣势 |
|---|---|---|
| 单个B细胞技术 | 1.高效率 2.抗体重轻链天然配对 3.无需培养B细胞 4.保留抗体多样性 |
技术门槛高 |
| 杂交瘤技术 | 抗体重轻链天然配对 |
1.许多物种没有可用的融合伴侣细胞 2.细胞融合效率低 3.抗体制备所需时间久 |
| 噬菌体展示技术 | 1.无物种限制 2.可通过控制筛选策略,选择所需特异性的抗体 |
1.抗体重轻链非天然配对 2.建库较为困难 |
免疫球蛋白A(IgA)是黏膜免疫系统中最主要的抗体成分,也是人体每天产生最多的抗体。与其他免疫球蛋白相比,IgA在分子形式、糖基化模式以及存在1个以上的受体等方面具有独特的性质。
在人体中,IgA单体以两个密切相关的亚型存在,即IgA1和IgA2,区别在于IgA1分子铰链区中多13个额外的氨基酸。这种延伸的铰链区有利于较远距离抗原的识别。两个IgA亚型在其CH1和CH2结构域中都存在高度N-糖基化,糖类约占其含量的6%。IgA1在铰链区含有额外的O-连接聚糖,由N-乙酰半乳糖胺与半乳糖和唾液酸组成。值得注意的是,IgA1铰链区的聚糖组成是异质性的,据报道异常糖基化与IgA肾病的发病机制有关。
IgA主要以三种不同的形式存在。在人体血清中以单体形式存在,而在粘膜部位,它以聚体的形式存在(pIgA),可分为二聚体IgA(dIgA)和分泌型IgA(sIgA)。

除少数例外,几乎所有治疗用的抗体均为IgG类,这可以归因于该抗体类别在人体内的天然丰度,长半衰期,但也归因于对IgG的全面了解和对其他同种型了解有限。
IgA作为治疗性抗体主要具有以下优势:与其他免疫球蛋白同种型相比,IgA可以与多种受体相互作用,包括多聚免疫球蛋白受体(pIgR)、转铁蛋白受体(CD71)、脱唾液酸糖蛋白受体(ASGPR)、dectin-1、DC-SIGN和FcαRⅠ(CD89)等,能更有效地激活细胞效应功能。以癌症研究背景为例,IgA可以通过FcαRⅠ诱导比IgG更强的中性粒细胞介导的肿瘤细胞杀伤力,相比之下IgG缺乏中性粒细胞的有效活化。这可能是由于IgA介导的LTB4释放导致中性粒细胞迁移增强。此外,IgA通过ADCC、氧化爆发(oxidative burst)、细胞因子和中性粒细胞的NETs释放诱导更强的吞噬作用。尽管FcαRⅠ表达低于FcγR,但IgA-FcαRⅠ结合诱导比IgG-FcγR结合更强的ITAM信号传导。

与IgG相比,IgA除了通过FcαRⅠ ITAM信号通路介导更强的免疫激活外,IgA还可以通过抑制性ITAMi途径介导更强的免疫调节作用。这些免疫调节特性在治疗炎症性疾病中具有很大的治疗潜力。
IgA的研究和临床使用可能由于以下几个原因受到阻碍:(1)小动物模型的问题,尤其是啮齿动物。啮齿动物与人类IgA有很大不同,小鼠和大鼠B细胞仅产生一个亚型的IgA,且其在血清中是以二聚体形式存在。存在两个亚型IgA这一特征仅与黑猩猩、大猩猩和长臂猿共享。主要的IgA受体FcαRⅠ在啮齿动物中缺失,并且人IgA在小鼠中只有很短的半衰期。(2)重组生产IgA的问题。由于IgA存在高度和异质的糖基化,生产过程中可能引起糖基化类型的改变,增强免疫原性和清除率。(3)与IgG相比,IgA半衰期短,需要较短的给药间隔。
多年来,sIgA一直被认为是对抗粘膜表面病原体的第一道屏障。sIgA能凝集细菌,干扰细菌运动,中和细菌毒素,还能抑制细菌对上皮的粘附,从而防止病原体向循环传播。已经证实IgA可对抗多种胃肠道病原体,例如鼠伤寒沙门氏菌,福氏志贺氏菌,艰难梭菌以及某些病毒。特别是,IgA对仙台病毒、人类免疫缺陷病毒和流感病毒具有中和作用。最近研究表明,IgA在中和SARS-CoV-2方面比IgG更有效。IgA还可以通过与FcαRⅠ的相互作用介导对微生物感染的保护。已经证明,在人FcαRⅠ转基因小鼠中输注抗原特异性IgA可以导致结核分枝杆菌或百日咳博德特氏菌的清除率增强。基于这些观察结果,被动转移特异性IgA和主动免疫可能是对抗病毒和细菌感染的有效策略。

IgA的治疗潜力
轮状病毒疫苗被视为了解肠道IgA在胃肠道病毒感染中的治疗潜力的模型系统。在该疫苗开发前后,几项相关研究表明,轮状病毒特异性IgA是赋予人类和动物模型长期免疫力的主要效应分子之一。目前的两种口服疫苗Rotarix®(葛兰素史克生物制品)和RotaTeq®(默克)于2006年获准使用。接种两剂疫苗后,血清抗轮状病毒IgA阳转率约为95%,可通过血清IgA滴度预测保护持续时间和疫苗效力。
原发性免疫球蛋白缺乏症(PAD)患者的免疫球蛋白水平降低,易发生感染。使用IgG替代疗法成功地降低了严重细菌感染的频率。然而,非呼吸道和上呼吸道感染持续存在,尤其是IgA和IgM水平低的患者。一项研究报告证实,使用Pentaglobin®(一种含IgA和IgM的制剂)成功治疗PAD患者的空肠弯曲菌复发性感染。与Pentaglobin®相比,Trimodulin(BT-588)的IgA含量其两倍,可限制重度社区获得性肺炎患者的继发感染。
值得注意的是,没有关于选择性IgA缺乏症治疗的报道(选择性IgA缺乏症是最常见的PAD)。这可能是由于对Ig产品的反应和抗IgA抗体的出现。过往曾出现缺乏IgA的患者由于输注IgG(含有少量IgA)而出现抗IgA抗体,导致全身性过敏反应。但也有综述详细描述了抗IgA抗体的产生与过敏反应的发生并不相关。未来,需要大型研究来评估含有IgA的产品在预防感染方面的安全性和治疗效果,特别是在选择性IgA缺乏症的治疗中。
用于治疗各种癌症的治疗性抗体通过直接和间接效应功能的组合来消除肿瘤细胞,包括补体依赖性细胞毒性(CDC)、抗体依赖性细胞吞噬作用(ADCP)和抗体依赖细胞介导的细胞毒作用(ADCC)。虽然IgG抗体在治疗领域占主导地位,但应该注意的是,IgA通过表达FcαRⅠ的巨噬细胞和中性粒细胞的活性也可以有效杀死肿瘤细胞。在这方面,体外实验结果表明,与IgG相比,IgA在中性粒细胞触发ADCC方面更胜一筹。随着FcαRⅠ转基因小鼠的产生,体内实验模型得到了极大的改善。在各种实体瘤模型中,已经证明抗EGFR IgA2抗体具有有效的抗肿瘤活性。在小鼠淋巴瘤模型中,抗CD20 IgA2在中性粒细胞募集到肿瘤部位后引起强大的抗肿瘤作用。
IgA抗体的半衰期很短,到目前为止阻碍了它们作为治疗性抗体的使用。最近的糖工程策略显著改善了重组IgA的药代动力学特性。例如,与野生型IgA相比,唾液酸化增加和聚糖末端半乳糖残基缺失的IgA分子表现出更长的血清半衰期,为免疫治疗提供了一种有希望的形式。另一种策略通过与可以结合白蛋白的结构域融合来增加IgA的半衰期。在人体中,可延长其血清半衰期至19天。除药代动力学外,IgA和CD47-SIRPα阻断剂的联用也提供了证据,表明靶向吞噬检查点抑制剂可增强IgA功能,这与IgG一致。
IgA与FcαRⅠ结合传播抑制信号,导致抗炎反应。因此,靶向FcαRⅠ可能是治疗各种炎症性疾病的一种有希望的策略。事实上,发现向FcαRⅠ转基因小鼠施用mIgA可预防和解决实验诱导的关节炎。同样,在类风湿关节炎患者中,mIgA能够在体外抑制促炎细胞因子的产生和骨髓细胞的趋化性。抗FcαRⅠ Fab片段可以驱动ITAMi诱导的抑制信号传导,并已在肾脏炎症模型中证明了其治疗潜力。还有研究表明,用抗FcαRⅠ预处理FcαRⅠ转基因小鼠可大大减少支气管炎症的发展。综上所述,FcαRⅠ靶向可能是预防或治疗炎症性疾病的一种新的有前途的工具。
在IgA半衰期和生产方面的研究进展为进一步拓展商业规模IgA生产提供了有意义的改善。由于IgA是对抗粘膜表面病原体的主要抗体,最近的研究还针对口服IgA进行了开发。全长sIgA或嵌合IgA已经被添加到食品中,用于在体外中和细菌毒素或预防胃肠道感染。未来的工作还应集中在简化制造工艺,降低生产成本上,也可汇集IgA和IgG同种型的优势,考虑将交叉同种型分子或双特异性抗体的工程设计用于治疗应用。
参考文献
[1]Sterlin D, Gorochov G. When Therapeutic IgA Antibodies Might Come of Age. Pharmacology. 2021;106(1-2):9-19. doi: 10.1159/000510251.
[2]Bohländer F. A new hope? Possibilities of therapeutic IgA antibodies in the treatment of inflammatory lung diseases. Front Immunol. 2023 Mar 27;14:1127339. doi: 10.3389/fimmu.2023.1127339.
[3]Breedveld A, van Egmond M. IgA and FcαRⅠ: Pathological Roles and Therapeutic Opportunities. Front Immunol. 2019 Mar 22;10:553. doi: 10.3389/fimmu.2019.00553.
重链和轻链异四聚体的IgG抗体形式曾是抗体结构的不二法则。直到Hamers-Casterman等人1993年在骆驼科物种单峰驼(Camelus dromedarius)中发现了一种新的抗体结构——重链抗体(HCAbs)。重链抗体的可变区结构域(VHH),又称纳米抗体,这种新型抗体的独特结构和新颖特性使其在各领域均有广泛的应用价值。
对骆驼科所有物种的免疫学调查研究显示,骆驼科动物体内均含有常规抗体(Conventional IgG)和HCAbs两种结构的抗体。常规抗体(IgG1)包含两条轻链(由VL和CL结构域组成)和两条重链(由VH、CH1、铰链、CH2和CH3结构域组成),两种重链抗体(IgG2和IgG3)为仅包含重链的同二聚体,每个重链包含VHH、铰链、CH2和CH3结构域。IgG2型的铰链长度较IgG3型长。

图1 骆驼血清中天然存在的抗体示意图
多项研究对骆驼科动物IgG1抗体VH序列的相似性、多样性以及CDR区长度分布等几个方面进行分析,证实骆驼科动物IgG1的VH序列与人抗具有高度一致性,且VH(4)家族CDR3区长度分布与VHH CDR3区类似,长于小鼠IgG。
骆驼科动物和人类VH序列具有高度的一致性。有研究发现,骆驼VH(3)共有序列与美洲驼框架区残基序列一致性可达100%,与人类框架区序列之间的一致性可达94%,只有残基49,74,83和84的4个保守差异。骆驼VH(4)共有序列与人类共有框架序列的同源性稍低,为86%。各种哺乳动物免疫球蛋白序列的框架一致区域的系统发育分析显示,单峰驼、美洲驼、双峰驼VH(3)序列聚类最接近人类VH(3);而单峰驼和双峰驼VH(4)序列与人类VH(4)、绵羊VH(2)和小鼠VH(2)均在同一聚类。

图2 骆驼科动物重链可变区框架序列与其他哺乳动物物种的相应区域间的系统发育树
可变区基因的体细胞高频突变增加了抗体多样性,特别是在CDR环中,在框架区中也是如此。有研究基于57个骆驼科IgG1抗体序列总结了其框架区的变异性(图3),结果表明:(1)在框架1和3中观察到的变异性高于框架2;(2)VH(3)框架的变异性始终高于VH(4)框架:这可能反映了VH种系基因使用的差异,以及随着免疫反应的成熟对VH(3)的偏好高于VH(4);(3)与IgG1a亚型相比,IgG1b亚型中框架区域的更大变异性,可能是由于免疫应答成熟过程中的同型转换。

图3 VH、VK和VL框架区序列变异性
在另一项研究中,研究者使用Mi-Seq系统对三头双峰驼体内抗体进行测序,测序结果表明:(1)框架区比CDR区相比更为保守,这与先前的认知一致;(2)框架2区不如其他框架区保守;(3)CDR2区的多样性高于所有其他区域;(4)VH和VHH的CDR3多样性是相似的。

图4A FR1, CDR1, FR2, CDR2, FR3, CDR3和FR4的子区域的唯一序列数量
图4B 归一化后的多样性评价,等于唯一序列数量除以每个子区域的平均序列长度
基于Kabat定义比较了骆驼科动物VH(3)和VH(4)序列之间的差异,图5ab详细描述了CDR-H1和CDR-H2长度的变异性。在34条VH(3)序列中,CDR-H1均为5个残基;在所有23条VH(4)序列中,CDR-H1除1条例外为6个残基,其他均为7个残基。VH(4) CDR-H2只出现包含16个残基一种可能性,而VH(3) CDR-H2的范围为16~22个残基,众数为17个残基。
和VH(4)序列CDR-H1、CDR-H2长度分布")
图5 骆驼科动物VH(3)和VH(4)序列CDR-H1、CDR-H2长度分布
CDR-H3观察到的长度变化最大。VH(3)序列通常显示较短的CDR-H3长度,残基数量范围3~18,其中最常见的情况是12~13个残基。相反,VH(4)序列的CDR-H3长度分布更集中,从11到20个氨基酸,最高频率的为14和17个氨基酸。
许多研究报道,骆驼科动物VHH结构的一个关键特征是较长的CDR-H3区。结构研究表明,VHH结构可以利用其较长的CDR-H3环更好的与目标蛋白表面缝隙(clefts)结合。将其与骆驼科动物的VH的CDR-H3长度进行比较发现,32条VHH CDR-H3序列中只有7条(22%)长度大于VH(4)的20个残基,所有VH(4) CDR-H3序列长度都在VHH抗体的CDR-H3长度范围内(7~24个残基);与公共数据库(http://www.bioinf.org.uk/abs)中提取的人类和小鼠VH CDR-H3长度进行比较,可以发现骆驼VH(3) CDR-H3长度分布与小鼠IgG相似,而VH(4) CDR-H3长度较小鼠IgG更长。由此我们可以进行推断,与VH(3)相比,VH(4)对较长CDR-H3长度的偏好是否也能使相应抗体更好的与蛋白表面结合。
和VH(4)序列CDR-H3长度分布")
图6 骆驼科动物VH(3)和VH(4)序列CDR-H3长度分布
参考文献
[1]Griffin LM, Snowden JR, Lawson AD, et al. Analysis of heavy and light chain sequences of conventional camelid antibodies from Camelus dromedarius and Camelus bactrianus species. J Immunol Methods. 2014 Mar;405:35-46.
[2] Li X, Duan X, Yang K, et al. Comparative Analysis of Immune Repertoires between Bactrian Camel’s Conventional and Heavy-Chain Antibodies. PLoS One. 2016 Sep 2;11(9):e0161801.
[3]Henry KA, van Faassen H, Harcus D, et al. Llama peripheral B-cell populations producing conventional and heavy chain-only IgG subtypes are phenotypically indistinguishable but immunogenetically distinct. Immunogenetics. 2019 Apr;71(4):307-320.
单克隆抗体(mAb)是制药市场上最重要的生物药物类型之一。目前,已有100多种单克隆抗体被批准用于治疗癌症、传染病、自身免疫性疾病和神经系统疾病等多种疾病。在当今快节奏和竞争激烈的药物开发市场中,加速开发单克隆抗体试剂是成功推进治疗性抗体的基本要求。
继经典的杂交瘤技术之后,已经开发和改进了许多其他方法,包括B细胞永生化技术、各类展示技术以改善单克隆抗体发现的途径。单B细胞抗体开发技术是近年来发展起来的新型单克隆抗体制备技术,该方法依赖于磁珠、流式、微流控等各类单细胞分离鉴定技术,结合PCR技术、DNA测序技术和抗体重组表达技术,实现从单个B淋巴细胞中分离表达天然配对的特异性抗体。单B细胞抗体开发技术实现了抗体的高通量筛选,克服了展示技术制备抗体的缺点,保留重链和轻链原始配对,能够筛选出高亲和力、特异异性的抗体。
该技术主要包括以下几个步骤:单个B细胞的分选、扩增和克隆抗体基因、抗体表达和鉴定。
B细胞亚群的选择
抗原特异性B细胞是获得抗原特异性单抗序列的主要来源,B细胞亚群的选择随技术路线的差异而不同。B细胞的发育是一个有序的过程,大致可分为祖B细胞(pro-B cells)、前B细胞(pre-B cells)、未成熟B细胞(immature B cells)、过渡B细胞(transitional B cells)和成熟B细胞(mature B cells)五个阶段。成熟B细胞到达外周免疫器官的B细胞区定居后,在那里接受外来抗原的刺激而活化、增殖进一步分化熟为浆细胞和记忆B细胞。

B细胞发育过程
浆细胞(plasma cell)是分泌抗体的终末分化细胞,具有非常低水平的膜免疫球蛋白表达,但表达MHC Ⅱ类分子(在分化过程中可通过负反馈途径抑制Tfh活化)。成熟B细胞除分化成浆细胞外,还有部分分化为记忆B细胞(memory B cell)。记忆B细胞表达膜免疫球蛋白但不分泌抗体。一般来说,记忆B细胞和浆细胞均在生发中心经过体细胞高频突变和亲和力成熟,其抗原亲和力较其他前体B细胞更佳,是单B细胞抗体开发常选择的B细胞亚群。
单个B细胞的分选方法
单个B细胞分选可分随机分离和抗原特异性分离,前者只需分离单个B细胞,操作较为简单,后者需分离抗原特异性B细胞,操作较复杂,尤其适合抗肿瘤抗体、自身免疫抗体等特异性抗体含量较低的情况。
随机B细胞分离的方法主要有显微操作法、激光捕获显微切法、流式细胞分选法等。抗原特异性B细胞分离的方法包括:磁珠分选法、荧光标记多参数流式细胞分选法、微流控分选法、微雕法以及细胞微阵列芯片法等。
磁珠分选法基本原理是基于B细胞表面标志物能与连接有磁珠的目标抗原相结合,在外加磁场中,通过与目标抗原结合的细胞被吸附而使细胞得以分离。这种方法可以在几分钟内从复杂的细胞混合物中分离出高纯度的细胞,在流式分选单细胞前可用磁珠分选法进行预分离,但是由于磁珠影响细胞生物活性因而不利于分离后培养与操作。
荧光标记多参数流式细胞分选法与流式细胞分选法类似,二者差异在于前者在加入用于标记B细胞表面标志物的荧光抗体之外还需加入荧光标记的目标抗原用来结合B细胞表面BCR以分选目标抗原特异性B细胞。该技术优点是可快速、准确、高通量分离B细胞并进行多参数同时分析。
微流控分选技术基于一个全封闭的检测系统。该技术可以将单个抗体分泌细胞与包被抗原的微球及捕获抗体的荧光标记二抗共同包裹在一个微液滴中。每一个微液滴相当于一个独立的微型反应器,抗体分泌细胞在液滴中培养一段时间后,通过荧光激活液滴分选系统可分选出含有目标细胞的液滴。
B细胞通常难以维持较长时间的离体存活,这对鉴定表达抗原特异性抗体的B细胞提出了挑战。可通过EBV转化、慢病毒介导的遗传修饰以及CD40L联合多种细胞因子的体外诱导培养等B细胞永生化技术延长细胞离体存活时间。
细胞分选时,通常需将单个B细胞分至内含适量细胞裂解液、RNA酶抑制剂和PCR反应试剂的适当容器中,如96孔板。由于单个细胞内RNA含量少,适当的容器可以方便大批量操作、防止样品损失或交叉污染。另外,不同类型B细胞抗体分泌能力差异明显,如抗体分泌B细胞中抗体基因转录本含量远高于记忆B细胞,因此从抗体分泌B细胞中更容易扩增得到抗体基因。
通常从单个B细胞中扩增未知抗体基因,需使用合适的引物进行巢式或半巢式逆转录PCR(Nested or semi-nested RT-PCR),该过程要求引物具有通用性、灵敏性、特异性,能避免非特异性扩增又能扩增出完整的抗体基因序列,因此合理设计引物序列至关重要。通常针对抗体重链轻链可变区不同前导序列设计前向引物的混合物,反向引物特异性互补于抗体恒定区。根据实验目的,如果分离和扩增不同同种型的抗体,反向引物则是特异性互补于各种同种型抗体恒定区的混合物。扩增出的抗体基因片段连接成scFv、scAb、Fab等形式,酶切将其构建到原核或者真核表达载体中。
鉴定抗体的抗原特异性和生物活性前需将携带有抗体基因的表达载体在相应系统中表达,常用的是真核表达系统尤其是哺乳动物细胞表达系统,更有利于抗体的加工修饰,其产物的生物活性可靠性更高。常用的有HEK293和CHO等细胞系。根据目的抗体及实验目的不同,所选取的鉴定及筛选方法也各不相同。可通过SDS-PAGE、Western blotting、ELISA等常规方法筛选和鉴定抗体,也可通过流式检测、免疫沉淀、体内药物活性测定等方法进一步测定抗体与抗原的特异性、亲和力以及中和活性等生物学特性。
优势
劣势
参考文献
[1]Tiller T, et al. Single B cell antibody technologies. N Biotechnol. 2008;25(5): 333-7.
[2]迟象阳.单个B细胞抗体制备技术及应用[J].生物工程学报, 2012, 28(6):651-660.
在过去的20年中,治疗性抗体已成为治疗恶性、感染性和自身免疫性疾病的重要选择。凭借高特异性、治疗功效、稳定性、溶解性、临床耐受性和相对便宜的生产成本,单克隆抗体在靶向治疗领域实现了飞跃。
免疫原性即生物制剂诱导体液或细胞介导的免疫反应的能力,长期以来一直是治疗性抗体的“致命弱点”。治疗时非人抗体通常会被患者的免疫系统识别为外来抗体,导致治疗性抗体或其衍生物的快速清除,限制了疗效。嵌合抗体通过使用人抗恒定区取代鼠抗恒定区,以尝试降低免疫原性。在CDR移植抗体中,可变区的人源化程度进一步增加,帮助免疫原性特征显著改善。然而人源化甚至完全人源抗体在患者中或多或少具有免疫原性的问题仍有待解释。
与天然人源抗体相比,大多数人源化抗体的CDR区表现出不同的结构折叠组合,这为诱导针对治疗性抗体的不良免疫应答提供了合理的解释。这些规范结构(canonical structures)被证明对于在人源化抗体时保持亲和力至关重要,也开辟了超级人源化的方法,可以将鼠抗与人抗规范结构进行匹配。基于抗体的疗法突破的一个重要驱动力是人类、啮齿动物和其他物种种系免疫球蛋白(Ig)重链和轻链(V,(D),J和C)基因的测序/表征。它揭示了Ig库的广泛多样性,并解释了其识别和结合几乎任何具有不同亲和力水平的抗原的能力。现在有许多产生人类治疗性抗体的抗体发现平台可用,包括转基因小鼠系统以及可以通过各种展示技术筛选的非免疫和合成人抗体库。
基于已发表的美洲驼和单峰驼体内成熟常规抗体的VH区域,研究人员注意到骆驼科动物与人类抗体序列高度的同源性。根据全基因组鸟枪数据库(WGS)和高通量基因组测序数据库(HTG)中骆驼和羊驼数据,各自框架FR1、FR2和FR3的一致性,CDR1和CDR2的长度,以及规范结构的预测结果表明,骆驼科动物和人类抗体V基因库具有序列同源性。规范结构预测基于Chothia规范,除了CDR的长度之外,还考虑了CDR和FRs中关键残基的存在。此外,比对表明,在不同的骆驼物种之间,IGHV、IGLV和IGKV基因是同源的(有时甚至是相同的),并且共享相同的V区家族和亚家族。
骆驼科动物IGHV基因库主要包括VH1、VH3、VHH3和VH4基因家族。除了IGHV1、IGHV3、IGHV4基因家族外,还存在属于IGHV5和IGHV7基因家族。所有五个V基因家族都与人类对应基因家族高度同源。IGHV1基于匹配的规范结构1-2(HCDR1和HCDR2)进行鉴定,这些结构与人类种系hIGHV1-1、1-2、1-3相关。IGHV3家族确定了2个亚家族,可以根据其CDR2残基组成将其细分为4组(a-d)。IGHV3两个亚家族分别与具有规范结构1-3和1-1的hIGHV3-23和IGHV3-66相关。VHH3是骆驼科所特有的,根据1-3和1-1的规范结构,可分为2个亚家族,一个亚家族与hIGHV3-30/hIGHV3-11相关,一个亚家族与hIGHV3-66相关。在IGHV4家族中,可以鉴定出两个亚家族:一个与hIGHV4-30-4相关,规范结构为3-1;另一个与hIGHV4-30-4具有相同的HCDR1长度,但HCDR2不同,这可能导致与人类胚系IGHV4对应的规范折叠不同。IGHV5和IGHV7家族与人类IGHV5-51和IGHV7-4基因具有相同的规范结构1-2。
骆驼科动物IGHV1和IGHV3与人类IGHV家族的FR一致性92%~95%,IGHV4一致度至少82%。骆驼科动物IGHV5和IGHV7的FR序列一致性较低,但仍有77%。进一步分析显示,VHH3和VH3基因分别包含FR2的亲水性氨基酸残基替换(F42/Y42, E49/Q49, R50/C50和F52/G52/L52)和疏水性氨基酸替换(V42/I42, G49, L50和W52/S52)。

图1 骆驼科动物IGHV框架区与人抗相似度比对
骆驼科动物IGLV1家族由2个与hIGLV1-47和hIGLV1-40相关的亚家族组成,分别包含规范结构13-7和14-7。发现的大多数骆驼科动物IGLV2家族与hIGLV2-18有关,规范结构为14-7。IGLV3包含两个亚家族:一个与hIGLV3-9更相关,而第二个可以进一步分为3个与人类IGLV3-25相关(两者都具有规范结构11-7)。IGLV4家族可分为5个亚家族,且均与hIGLV4-60最相似,规范结构为14-11。IGLV5家族中10个亚家族与hIGLV5-39,hIGLV5-52,hIGLV5-37,hIGLV5-45一致性较高,所有亚家族(亚家族8除外)共享相同的14-11规范结构。IGLV6基因与hIGLV6-57相似,由于过早终止密码子,无法确定规范结构。IGLV7基因仅在骆驼和羊驼基因组中鉴定出,并被证明与hIGLV7-46相关,规范结构为14-7。IGLV8和IGLV9分别由8个亚家族和2个亚家族组成,分别与hIGLV8-61和hIGLV9-49相关,规范结构分别为14-7和12-12。IGLV10家族与hIGLV10-54相关,具有相同的规范结构13-7。
三种主要的人类IGLV家族(1-3)与骆驼科动物IGLV高度同源,基于FR的序列同一性在81%~91%。有趣的是,尽管总体一致性在61%到91%之间,但所有规范结构都与人类V区家族相匹配。此外,关键位点残基保守不仅局限于框架区,而且还扩展到CDR区,骆驼和人类V区基因之间框架区的这种高序列和结构相似性在免疫原性方面可能非常重要。

图2 骆驼科动物IGLV框架区与人抗相似度比对
骆驼科IGKV库由6个家族组成,骆驼科IGKV1家族拥有三个亚家族,与hIGKV1-27和hIGKV1-39相关,并具有相同的规范结构2-1。IGKV2家族拥有两个亚家族,与hIGKV2-29和hIGKV2-40相关,分别具有规范结构4-1和3-1。IGKV3和IGKV4亚家族分别与hIGKV3-11、hIGKV4-1相关,分别具有规范结构2-1和3-1。IGKV5和IGKV6家族与hIGKV5-1和hIGKV6-41相关,均基于2-1规范结构。
骆驼科IGKV基因与人类基因的同一性百分比在60%至85%之间,其中IGKV3基因的相似性最低。除了IGKV5,鼠IGKV基因与人类基因的亲缘关系更密切。同时,一致性不仅限于框架区,CDR区也是如此。

图3 骆驼科动物IGKV框架区与人抗相似度比对
为了进一步证实骆驼科动物抗体和人类抗体V区之间结构同源性,研究人员通过X射线晶体学确定两种骆驼科抗体的结构。这两种抗体是从使用人CD70或MET过表达细胞主动免疫的美洲驼制成的噬菌体展示抗体库中分离的。CD70(一种膜结合细胞因子)和MET(一种酪氨酸激酶受体)在肿瘤生长/存活、血管生成和转移中起重要作用。通过对获得的美洲驼抗体X射线晶体学进行结构测定,抗CD70(PDB 4R90; VH3/Vλ7)和抗MET(PDB 4R96; VH1/Vκ4)抗体结构分析见图4。
如图4C和4D,美洲驼抗MET(VH1/ Vκ4)单抗的环状结构与人类的重链和轻链(以PDB结构1FVD和1HIL为模板)相似。模板1FVD Fab中的HCDR1和HCDR2的规范结构1-2与抗MET Fab(属于IGHV1家族)的规范结构非常吻合,这意味着它确实利用了与匹配的人类种系VH1片段相同的规范折叠组合。此外,在1HIL模板中,LCDR1,LCDR2和LCDR3(灰色)的规范结构3-1-1分别与抗MET抗体的Vκ片段(蓝色)完美重叠。这证实了美洲驼衍生的IGKV4与人类IGKV4片段使用相同的规范折叠。总之,结构数据明确地证实了在骆驼科来源抗体的可变区域中存在的人类抗体的规范折叠。

图4 美洲驼重链和轻链V域和对应人抗的晶体结构
(A)美洲驼抗CD70抗体 VH(黄色核心,红色H1和H2,橙色CDR3)与人抗(1MFA,灰色)叠加效果图
(B)美洲驼抗CD70抗体 VL(粉色核心,绿色H1-H3)与人抗(灰色)叠加效果图
(C)美洲驼抗MET抗体 VH(绿色核心,蓝色H1-H2,红色CDR3)与人重链参考物1FVD(Vκ1)和1HIL(Vκ4)(灰色)叠加效果图
(D)美洲驼抗MET抗体 VK(绿色核心,蓝色H1-H3)与人重链参考物1HIL(Vκ4)(灰色)叠加效果图
参考文献
Alex Klarenbeek, Khalil El Mazouari, Aline Desmyter, et al(2015) Camelid Ig V genes reveal significant human homology not seen in therapeutic target genes, providing for a powerful therapeutic antibody platform, mAbs, 7:4, 693-706, DOI: 10.1080/19420862.2015.1046648
目前,已有数百种治疗性IgG抗体进入临床试验,90多种基于该同种型的抗体产品已获得FDA批准。事实上,IgM也已经在各种动物模型,包括非人灵长类动物中被证实是有治疗效果的,并已有首批IgM单克隆抗体药物进入了临床试验阶段。约20种IgM抗体在人体中进行了试验,包括针对各种传染病、肿瘤和自身免疫性疾病抗原的大鼠、小鼠和人源IgM。

图1 IgM抗体药物的临床试验史
截止现在,尚未有治疗性IgM抗体药物获批。这一结果可能是由于大多数进入临床的IgM抗体是天然来源,未经历广泛的体细胞突变,因此亲和力和特异性较低。药物早期研究中主要关注特定的适应症,如脓毒症和脓毒性休克,也可能限制了IgM抗体获得药物监管认可的可能。
表1 临床试验中的IgM抗体药物
| Antibody (Name) | Company | IgM Source | Antigen | Indication | Most Advanced Clinical Development |
|---|---|---|---|---|---|
| Campath-1M | Academic (MRC-RDCT) |
Rat | CD52 | Graft vs. host disease | Phase 2 |
| E5 (Xomen-E5) | XOMA | Mouse | J5 lipid A | Sepsis | Phase 3 |
| HA-1A (Centoxin) | Centocor | Human | J5 lipid A | Sepsis | Phase 3 |
| Fanolesomab-Tc99 (NeutroSpec) |
Palatine | Mouse | CD15 | Appendicitis | Phase 3 |
| IgM cocktail (5) | Cutter/Miles | Human | LPS | Sepsis | Phase 1 |
| Mab 16.88 | Academic (Free University Hospital) |
Human | Colon cancer antigen |
Colorectal cancer | Phase 1 |
| MAB-T88 | Chiron | Human | LPS | Neutropenia | Phase 1 |
| PAT-SC1 | Partridge | Human | CD55 isoform |
Gastric cancer | Phase 1 |
| ABX-CBL | Abgenix | Mouse | CD147 | Graft vs. host disease |
Phase 2/3 |
| L612 | Chugai | Human | Ganglioside GM3 |
Melanoma | Phase 1 |
| MORAb-028 | Morphotek/Eisai | Human | Ganglioside GD2 |
Melanoma | Phase 1 |
| AR-101 | Aridis | Human | LPS | Nosocomial P. a. pneumonia |
Phase 2a |
| mAb216 | Academic (Stanford) |
Human | CDIM | B-lineage ALL | Phase 1 |
| PAT-SM6 | Partridge | Human | GRP78 | Multiple myeloma | Phase 1/2a |
| ARG098 | Argenes | Mouse/Human (chimeric) |
FAS | Rheumatoid arthritis |
Phase 1/2 |
| rHIgM22 | Grant | Human | CNS myelin | Multiple sclerosis/neuronal degeneration |
Phase 1 |
| TOL101 | Tolerates | Mouse | ab TCR | Renal transplant | Phase 2 |
1964年,Barth及其同事发表了首个研究未进行改造的人IgM抗体在人体中药物代谢动力学(PK)的文章。测试使用的IgM抗体从健康人体血清中纯化,用碘-131放射性标记后,注射到七个正常成年人中。每天收集血清样本并在伽马计数仪中进行分析。研究发现,正常人IgM在人体中的消除半衰期的范围为3.8至6.5天,平均为5.1天。值得注意的是,人IgM的半衰期比人类中通常报告的人IgG半衰期(18~21天)低3倍,反映了IgM抗体不与FcRn受体结合的事实。
跟据几种治疗性IgM单抗的临床药代动力学试验结果,这些IgM在人体中的半衰期比Barth等人所描述的半衰期短。应该注意的是,这些实验之间存在几个关键差异。首先,Barth测试的抗体是正常人的IgM,因此它不会与人类抗原结合,而随后测试的许多IgM是会与人类抗原结合的。因此,经过临床测试的IgM与表达这些靶标的组织结合后,可能会更快地被清除。其次,Barth测试的材料是从人血清中分离出来的,而大多数其他IgM是在小鼠、大鼠或仓鼠(例如CHO)细胞中产生的。IgG的生产宿主细胞和培养条件的变化会导致糖基化的变化,IgM抗体也有类似的变化,因此PK实验的差异是在预料之中的。分析技术(同位素与ELISA)和受试者群体(正常与患病)也是差异的促成因素。
已有针对一系列适应症的啮齿动物或人IgM抗体药物进行了临床试验。在这些试验中,近400名受试者接受了高达27mg/kg的治疗剂量,并且没有报告明显的安全问题。重要的是,对于用人IgM抗体药物进行的临床研究,很少或未观察到免疫反应。(所使用的免疫原性测定的细节及其敏感性相对性暂时没有具体指明。)
几种人IgM抗体在临床上被安全使用令人鼓舞。理论上,IgM的多价性引发的抗体高亲合力可能导致药物脱靶结合,引发意想不到的毒性或药物的快速清除。一些靶向肿瘤靶标的天然分离的IgM抗体中,可能存在低亲和力(affinity)高亲合力(avidity)的结合,这可能导致药物脱靶。在迄今为止报告的临床研究中,这样的担忧并没有实际发生。后续这些问题只能通过进一步开发和临床试验其他IgM抗体候选产品来解决。
随着我们对各个蛋白表达系统和IgM抗体生产理解的不断深入,未来以IgM为原型开发的工程抗体,将会被用于治疗各种适应症。
IgM具有10或12个抗原结合位点,能够以高亲合力结合抗原靶标,对于在癌细胞或其他靶细胞上重复表达的细胞表面靶标,每个IgM分子可以多次参与抗原结合。此外,IgM结构的灵活性可以为在细胞表面上结合多个靶标提供适当的架构。因此,IgM特别适合靶向困难的抗原。在一些早期基于IgM的开发工作中,在临床试验中评估了针对肿瘤抗原中糖类部分或糖脂部分的抗体。在这些情况下,相应的IgG抗体对糖表位的亲和力可能不足以有效靶向,而IgM表现出适合生物治疗用途的强结合和效应功能。所选肿瘤靶点的另一个具有挑战性的场景是其在肿瘤上的低表达,特别是治疗抵抗性肿瘤。基于IgM的高亲合力抗肿瘤药物可能对低表达或其他困难的靶标产生更加有效的抗体。
许多早期IgM人体临床试验中使用的是天然IgM抗体,这些抗体通常从患者体内分离出来或从小鼠杂交瘤中人源化获得。未来,工程化的IgM抗体存在着巨大的可能。如将亲和力成熟后的IgG抗体可变区嫁接到IgM恒定区上,这种设计将帮助IgM的结合亲和力和效应功能显著增加。除了结构域的嵌合外,也有研发平台试图通过IgG的工程改造,进一步利用IgM的六聚体结构。例如HexaBody™抗体平台是通过在IgG重链中引入突变,使突变后的IgG单体在特定浓度下的细胞表面以六聚体的形式存在。这种形式的抗体在保留ADCC等效应的同时,可以激发更强的CDC效应,综合了IgG和IgM效应功能优势,在治疗效果上较普通同种型更优。
我们还可以通过在IgM上添加多个特异性结合位点来优化抗体药物治疗功能。例如,双特异性IgG抗体和双特异性IgG抗体变体表现出极强的肿瘤靶向作用。然而,这些抗体只有一个与肿瘤抗原的结合位点。双特异性IgM或许可以与困难或罕见的肿瘤抗原建立非常高的亲合力结合,选择性地结合T细胞以实现有效的肿瘤细胞杀伤。例如,一类将CD3结合位点与J链融合来产生T细胞效应功能的双特异性IgM抗体,该抗体包含10个TAA结合位点和CD3结合位点。这种抗体结构的一个关键特征是能够在单个高表达细胞系中制造完全组装的双特异性IgM抗体。
随着对IgM抗体和IgM抗体工程的重新关注,IgM平台很可能凭借其固有优势,产生改进的生物治疗制品,用于治疗未满足的医疗需求。我们预计未来具有增强受体交联的IgM的高阶价态和高效的双特异性IgM应为抗体工程和开发更有效的治疗方法提供新的机会。
参考文献
Keyt BA, Baliga R, Sinclair AM, et al. Structure, Function, and Therapeutic Use of IgM Antibodies. Antibodies[J], 2020, 9(4): 53. doi:10.3390/antib9040053
细胞毒性T细胞是癌症患者治疗过程中效力最高的免疫细胞群。1992年,重组IL-2(一种T细胞活化细胞因子)获得批准,以及T细胞检查点抑制剂(如PD-1拮抗抗体帕博利珠单抗和纳武利尤单抗)的临床影响,获得了T细胞在癌症治疗中重要性的早期临床证据。更直接的证据来自自体肿瘤浸润T细胞(TILs)的治疗方法,该疗法与清淋治疗和IL-2相结合,在黑色素瘤和卵巢癌患者中产生了强烈的反应。当自体T细胞被改造为表达重组嵌合抗原受体(CAR)时,只要它们能够对细胞表面抗原进行主要组织相容性复合体(MHC)非依赖性识别,就可以在液体肿瘤中观察到高完全缓解率和持久反应,已有几种靶向CD3或BCMA的CAR-T疗法获得批准。尽管细胞疗法在非实体瘤中获得临床成功,但该治疗可能具有显著毒性,且在实体瘤中的活性有限。
T细胞衔接器(T cell engager, TCE)是细胞因子、检查点抑制剂和T细胞疗法的替代疗法,它们通常基于抗体或抗体片段,能够将体内任何T细胞与表达选定表面标志物的细胞连接起来。TCE可以精确控制剂量和摄入时间,有助于最大限度地减少副作用,并且生产过程比细胞疗法要简单得多。靶向CD19、CD20、BCMA的TCE在复发/难治性(r/r)急性淋巴细胞白血病(ALL)、非霍奇金淋巴瘤(NHL)、多发性骨髓瘤(MM)已经观察到令人信服的完全缓解率。
TCE一个臂靶向肿瘤相关细胞表面抗原(TAA),另一个臂靶向TCR-CD3复合物中CD3ε亚基,结合癌症细胞和T细胞。这种结合促进T细胞和癌细胞之间免疫突触的形成,T细胞可以通过突触将穿孔素和颗粒酶注入到附着的癌细胞中。突触的形成引起TCR交联和T细胞活化,引发促炎细胞因子的释放,诱导T细胞增殖。从理论上讲,每种细胞毒性T细胞表型都可以通过这种方式结合并参与连续癌细胞裂解,包括CD8+T细胞,CD4+T细胞,γ/δT细胞和NKT细胞。通过识别细胞表面抗原,TCE使得T细胞的活化摆脱了MHC分子、抗原肽加工和呈递以及TCR特异性的限制。

IgG-like TCE
Mosunetuzumab采用knobs-into-holes技术进行重链配对,采用体外退火技术进行重轻链配对,已被批准用于治疗R/R滤泡性淋巴瘤(FL)。

Teclistamab是一种基于Genmab公司开发DuoBody®平台的TCE,靶向BCMA/CD3,已上市用于治疗r/r多发性骨髓瘤(MM)。
DuoBody®平台利用限制性Fab臂交换技术(controlled Fab-arm exchange, cFAE),在两个IgG抗体的CH3区域分别引入K409R和F405L突变,利用哺乳动物细胞表达系统进行单独的重组表达与纯化,随后将两种抗体在定制条件下利用cFAE重新组合,即可高效组装获得具有正常IgG结构的双特异性抗体。Teclistamab抗体骨架为IgG4亚型,需要对其进行PAA突变(S228P L234A/L235A)除去IgG4的天然FAE过程和Fc的相关响应功能(ADCC,CDC)。

Epcoritamab也基于DuoBody®平台,最近被批准用于治疗R/R弥漫性大B细胞淋巴瘤(DLBCL)。
ERY974是一种靶向Glypican-3(GPC3)的TCE,用于治疗实体瘤,目前处于临床前开发阶段。它的构建是通过在CH3结构域引入静电转向突变来驱动重链的异源二聚化,同时利用共同轻链来避免轻链的错配(ART-Ig)。
REGN1979为靶向CD20/CD3的TCE,该TCE使用共同轻链,并在重链CH3区域中引入突变,形成IgG1/IgG2/IgG4与IgG3的局部同型嵌合体,利用蛋白质A的选择性纯化获得双特异性抗体。REGN1979现正在适应症恶性淋巴瘤(B-NHL)的临床试验中。

这种TCE结构有望成为一种新平台。基于该平台,还有一些用于治疗实体瘤的TCE,包括REGN4018——靶向MUC16/CD3,用于治疗卵巢癌(OC)和REGN4336——靶向PSMA/CD3,用于治疗转移性去势抵抗性前列腺癌(mCRPC),正在临床试验中。
基于抗体片段的TCE
虽然Fc-like TCE具有天然IgG的特性,有利于抗体药物的制备和PK/ADA特性,但或出于独特的医疗目的,或由于生产工艺技术或商业限制,基于抗体片段的TCE也逐渐走入视线。
基于抗体片段的TCE通常是——
这类TCE的代表是Amgen公司基于BiTE®平台,靶向CD19/CD3的Blinatumomab,已被批准用于治疗成人急性淋巴细胞白血病(BCP-ALL)及后续更多的适应症。
为了延长半衰期,减少给药频率,Amgen在BiTE®上融合Fc结构域,产生了一系列HLE BiTE分子(分子量约为106 kD)。AMG757是一种靶向DLL3的HLE BiTEs,目前正在研究用于复发性小细胞肺癌(SCLC)治疗。
以类似的方式,MacroGenics开创了DART平台并以此平台设计了治疗性分子,这些分子将两个独立的fab以异源二聚体结构结合在一起,可根据半衰期长短进行调整,具有或不具有Fc结构域。MGD024是一种用于治疗r/r CD123(+)血液系统恶性肿瘤患者的靶向CD123/CD3的Dart TCE,目前已进入临床研究。

Harpoon公司开发了针对实体瘤和血液系统恶性肿瘤T细胞激活性多功能抗体,抗体基于其创建的TriTAC、ProTriTAC(前药)和TriTAC-XR三个三特异抗体平台,这些TCE平台引入了单域抗体,并通过结合白蛋白来延长半衰期。
TriTAC仅含有一条肽链,包含三个结构域,分别结合CD3,人血清白蛋白和TAA。其中结合CD3表位的抗体为单链抗体(scFv),结合人血清白蛋白和TAA的抗体为纳米抗体。
此外,Immunocore公司开发的第一种也是唯一一种被批准用于实体瘤(黏膜黑色素瘤,mUM)治疗的TCE——Tebentafusp-tebn(KIMMTRAK),由TCRm和靶向CD3的scFv组成。

其他类型的TCE
EMB-07是利用EpimAb的FIT-Ig平台设计的,靶向ROR1/CD3的TCE,用于实体瘤的治疗,目前正在进行临床评估。
Appended-IgG在常规IgG上附加scFv、Fab或纳米抗体。它们的半衰期与亲代IgG相当。然而,非自然结构可能会增加聚集性、不稳定性和ADA风险。FIT-Ig平台在不引入突变或连接子(linker)的情况下产生双特异性抗体,有利于其制备和稳定性。
EMB-06是另一种基于FIT-Ig平台的靶向BCMA的TCE,目前也在临床评估中。
BA1202为Boan Biotech靶向CEA/CD3的对称三价单链抗体,其具有类似蝴蝶形的抗体结构:Fab端与CEA双价结合,而轻链C端连接的二价单链抗体与CD3单价结合(可能是由于空间构象的改变或亲和力降低),目前已进入临床试验阶段。
Cibisatamab是一种靶向CEA/CD3的TCE,通过将第二个与CEA结合的Fab区添加到对称KIH结构来实现与疾病靶点的“低亲和力(affinity)高亲合力(avidity)”结合,试图通过减少脱靶毒性来实现疗效和安全性的平衡。目前该TCE正在临床试验中。

CX-904是由Cytomx公司为精准靶向设计的前体药物,在肿瘤微环境中通过特异性蛋白水解条件被激活,靶向EGFR/CD3,目前正在临床试验中。
Imvotamab是靶向CD20和CD3的IgM同种型TCE。Imvotamab拥有10个CD20结合域,比IgG同种型的抗体有更高的亲合力,从而能够克服靶点减少引起的耐药,并实现比目前已批准的抗体疗法更好的疗效。该公司更多的IgM TCE药物包括IgM-2644(CD38/CD3)和IgM-2537(CD123/CD3)。
TNB-486是一种由Tenobio公司开发靶向CD19的不对称TCE。它的形式介于IgG-like TCE和基于抗体片段TCE之间。它由一半结合CD3的IgG和一半结合CD19的重链抗体(HcAb)组成,目前正针对r/r B淋巴瘤进行临床评估。
与TNB-486格式相似,YZYBio公司的YBODY平台由一半与疾病靶点结合的IgG和一半与CD3结合的scFv组成。一些基于YBODY平台的TCE目前正在临床试验中,如M802(HER2/CD3)。

参考文献
[1]Baeuerle PA, Wesche H. T-cell-engaging antibodies for the treatment of solid tumors: challenges and opportunities. Curr Opin Oncol. 2022 Sep 1;34(5):552-558. doi: 10.1097/CCO.0000000000000869.
[2]Pillarisetti K, Powers G, Luistro L, et al.Teclistamab is an active T cell-redirecting bispecific antibody against B-cell maturation antigen for multiple myeloma. Blood Adv. 2020 Sep 22;4(18):4538-4549. doi: 10.1182/bloodadvances.2020002393.
[3]Smith, E., Olson, K., Haber, L. et al. A novel, native-format bispecific antibody triggering T-cell killing of B-cells is robustly active in mouse tumor models and cynomolgus monkeys. Sci Rep 5, 17943 (2016). https://doi.org/10.1038/srep17943.






南京德泰生物工程有限公司 Nanjing Detai Bioengineering Co.,Ltd. ©2026 All Rights Reserved
 苏公网安备32011202001300
苏公网安备32011202001300